The main models of associations of GCA with cortical characteristics, and their change, were run separately for samples within the Lifebrain consortium (n = 1129, 2606 scans)20 and the UK Biobank (UKB, n = 2198, 4396 scans)21,22, and then meta-analyses were run on the results, using the metafor package26. Using the estimate and standard error at each vertex, random effects meta-analyses were conducted at each vertex separately. In all main models, sex, baseline age, scanner, time (interval from baseline) and education were entered as covariates. In modeling the effects of GCA on cortical characteristics (level-level analyses), GCA was entered as the predictor (explanatory variable), whereas in modeling the effects of GCA on brain aging (level-change analyses), the interaction term of GCA × time was entered as the predictor, and education × time was entered as an additional covariate along with GCA and education. Since brain aging (i.e. change) was of chief interest, we did not include intracranial volume (ICV), which is stable, in the main analyses. For direct comparison, we also then present level-level analyses without controlling for ICV. This was also chosen given the paucity of evidence for region-specific associations, and previous studies indicating that neuroanatomical volume in and of itself, when controlling for sex, may be associated with GCA9,25. Results from models including ICV, as well as models without education, as covariates, can be found in the Supplementary Information (SI). Additional analyses included the interaction term baseline age × time as a covariate, and in one set of analyses we entered the interaction term baseline age × time × GCA as predictor (with relevant two-way interaction terms as covariates), to test if effects differ reliably across the lifespan.
GCA level: brain level analyses
Cluster p-value maps across Lifebrain and UKB for the relationship of GCA and cortical characteristics controlled for education, are shown in Fig. 1. For cortical volume and area, there were widespread positive associations of GCA bilaterally across the cortical mantle seen in all lobes. For area, significant effects were seen across 47.6% and 44.2% of the left and right hemisphere surface, respectively. For volume, similar numbers were 37.4% and 19.5% for left and right, respectively.
For cortical thickness, only minor positive effects were seen, in proximity of the left central sulcus, covering only 1.1% of the surface.
Results of analyses per sample, controlling and not controlling for education are shown in Supplementary Figs. 1 and 2. Effects were largely similar, though slightly more restricted spatially, when controlling, than when not controlling for education. When adding ICV as a covariate, the intercept effects for cortical volume and area in the meta-analysis shown in Fig. 1 became non-significant, with only a very small effect on cortical thickness in the left hemisphere remaining (see Supplementary Fig. 3), pointing to these being broad effects grounded in greater neuroanatomical structures in general, rather than being region-specific.
To show effect sizes, we calculated the effect of 1 SD increase in GCA on cortical volume. Across Lifebrain and UKB, 1 SD higher GCA was associated with 1.0% larger cortical volume. Effect size maps for level-level analyses showing the regional variation in effect sizes for each sample separately are shown in Supplementary Fig. 4. Effect sizes were numerically smaller in Lifebrain (0.6%) than in UKB (1.3%). Restricting the analyses to regions where significant effects were seen, 1 SD increase in GCA was associated with 2.0% larger cortical volume in Lifebrain and 1.6% larger volume in UKB, but please note that these latter effect sizes are inflated by being within significant regions. Similar analyses for cortical area showed that 0.8% larger area was associated with 1 SD higher GCA across the cortex, with effects being 0.6% in Lifebrain and 0.9% in UKB. Restricting the analyses to regions where significant effects were seen, 1 SD increase in GCA was associated with 1.6% larger cortical volume in Lifebrain and 1.2% larger volume in UKB, with the same caveat as above. For thickness, effects were minute: 0.06% across studies (Lifebrain 0.04%; UKB 0.08%). Within significant clusters (UKB only), the effects of 1 SD higher GCA was 0.8%.
GCA level: brain change analyses
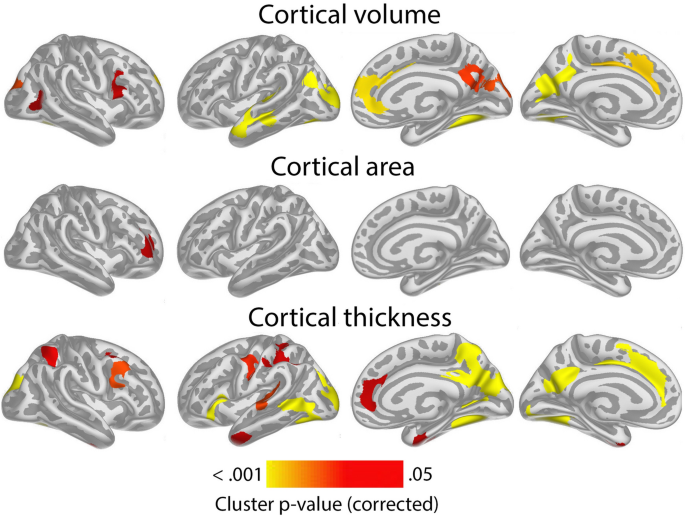
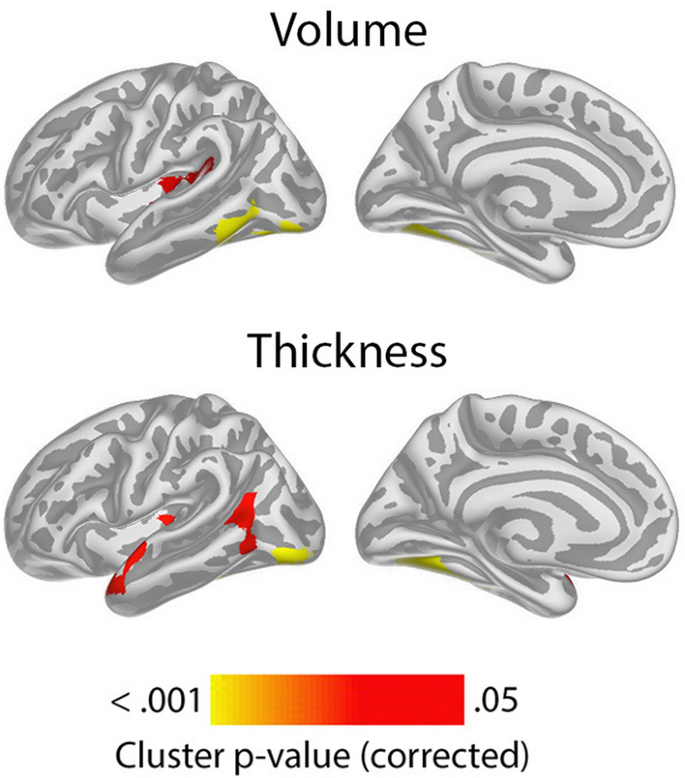
Having confirmed the expected positive relationships between GCA and cortical volume and area controlled for education in terms of an intercept effect, we investigated the question of slope effects. Associations of GCA level at baseline and change in cortical characteristics, controlled for education, are shown in Fig. 2. As expected, effects were more spatially limited than those seen for intercept models, with only restricted regions showing significant relationships: Higher baseline GCA was associated with less regional cortical volume reduction in the left middle cingulate gyrus, a medial area around the central sulcus and a part of the lingual gyrus. The most extensive effects were seen for thickness change, where higher baseline GCA was associated with less thinning in regions corresponding to the volume effects, in addition to parts of the right anterior and lateral temporal cortex and an area in the most medial part of the intersection between the central sulcus and the superior frontal cortex. No associations with area change were observed Taken together, this means that the observed positive associations of GCA with volume change primarily reflect less cortical thinning with higher GCA. (See Supplementary Fig. 5 for result for each subsample separately).
Associations of GCA with cortical change were essentially unaffected by adding ICV as a covariate (Supplementary Fig. 6).
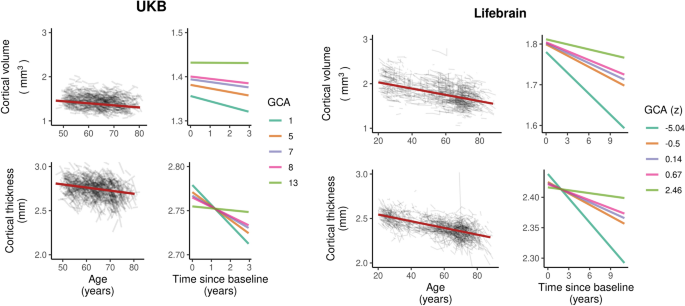
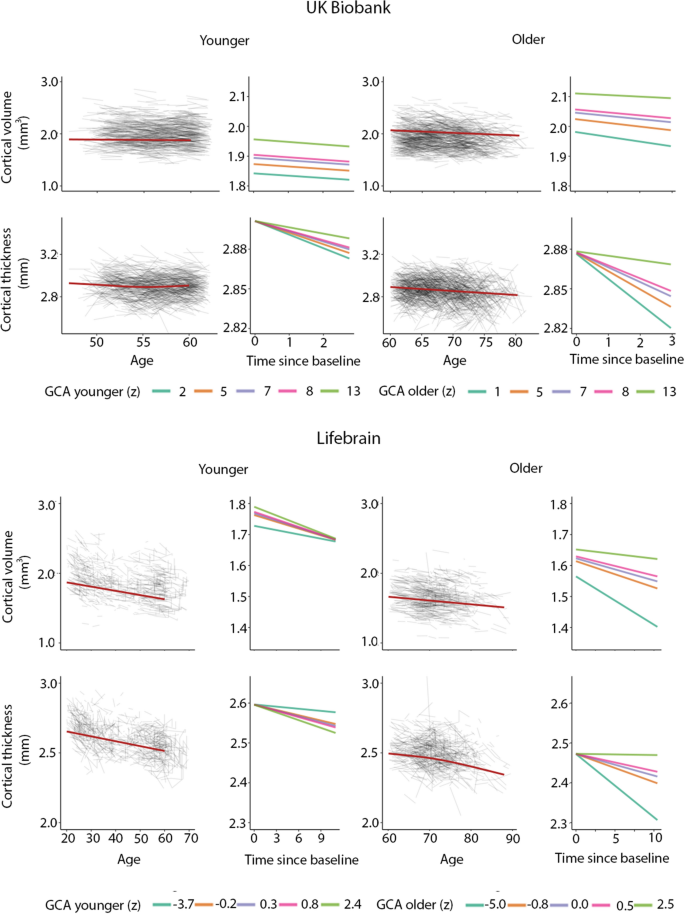
In order to illustrate the GCA-cortical change relationships, and to characterize consistency of effects across samples (UKB and Lifebrain), we plotted the generalized additive mixed model (GAMM) for the different GCA quintiles, from lowest to highest (Fig. 3), depicting change trajectories for average cortical volume and thickness within the regions showing significant GCA x time associations. Across samples, subgroups with higher GCA started with higher volume and had less volume loss over time. For instance, on average, people with maximum cognitive score in UKB are expected start out with a regional average cortical volume of 1.72 mm3 that would be maintained for the next three years, whereas those with the lowest GCA would on average start out with 1.64 mm3 and decrease to 1.61 mm3 over the next three years. Thus, the greatest GCA-associated differences in cortical volume are found in the intercepts (level), whereas differences in slope (change) are smaller in the follow-up period. For cortical thickness, the change trajectories were also very consistently ordered, but those with higher GCA did not uniformly have thicker cortex at first timepoint in these areas. Rather, differential rates of cortical thinning over time were critical in creating cortical thickness differences in these regions in aging. This was evident in both samples, but especially pronounced in UKB.
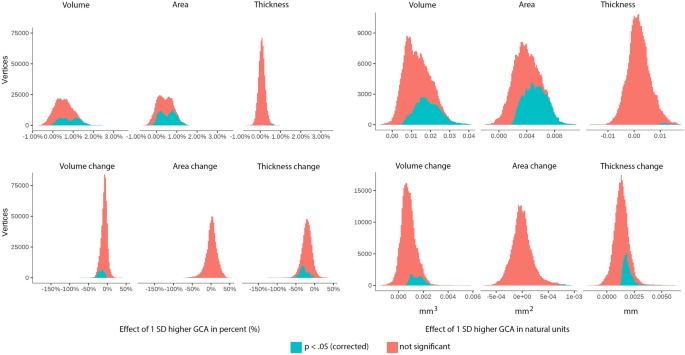
To further assess effect sizes, we created histograms of the vertex-wise distribution of effects of one SD higher GCA for each metric for absolute volume, area and thickness as well as their change, shown in Fig. 4. (For cortical distributions of such effect sizes per sample, see Supplementary Fig. 7). As can be seen, almost all vertices show positive level–level relationships between GCA and volume and area. For thickness, the distribution is only slightly shifted to the right of zero, confirming the weak GCA-thickness relationships. As for GCA level-brain change, the histograms showed that for area, effects were distributed almost perfectly around zero. For volume, there was a clear shift rightwards, meaning that higher GCA tended to be related to less volume reductions, but substantially less than for the offset effects. Cortical thickness showed the most rightward skewness of the distribution, much larger than for the offset results. Inspecting all histograms, it is clear that higher GCA is related to larger cortical volume and area, and less thickness change.
Influence of polygenic scores (PGSs) for GCA and education on the level-level and level-change associations
Next, we investigated whether effects were maintained when covarying for established PGSs for GCA and education in the UKB23,24. Fifty-two participants in the main models were excluded due to missing genetic data. In these analyses, we regressed out the first ten genetic ancestry factors (GAFs) from the GCA variable prior to analysis. The intercept associations of GCA and cortical characteristics that were observed in the main model (Fig. 1) largely remained when controlling for the PGSs, but the extent of the significant regions were somewhat reduced for cortical volume and area (Supplementary Fig. 8). The associations of GCA and cortical change largely remained and were only slightly attenuated when controlling for PGSs for GCA and education (Fig. 5; compare to UKB results in Supplementary Fig. 5).
Influence of age on the level-change associations
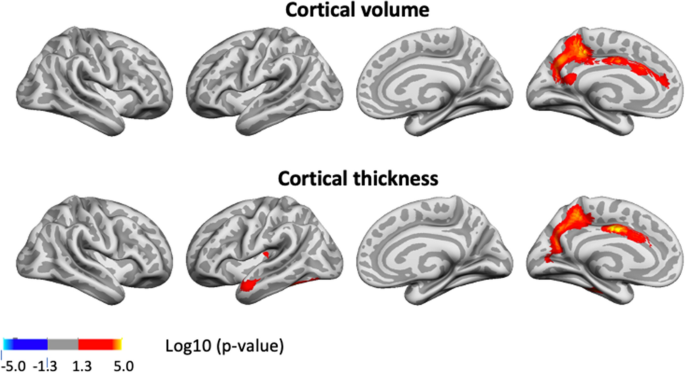
We next tested the three-way interaction baseline GCA × baseline age × time, to see whether the level-change associations differed reliably across the lifespan. Significant interaction effects were seen for change in small regions of the left hemisphere, mostly laterally for volume, and for slightly more extended regions, for cortical thickness (see Fig. 6).
The positive three-way interactions of baseline GCA × baseline age × time indicates that higher level of GCA is associated with less atrophy at distinct ages. To visualize these interaction effects, we divided the cohorts according to whether participants were above or below age 60 years. This division point was chosen in view of it being an approximate age at which select cognitive and regional cortical volume and thickness changes have been reported to accelerate in longitudinal studies27,28. In order to explore how GCA level related to cortical thickness change over time across the two age groups, we plotted the expected cortical change trajectories, within the significant regions shown in Fig. 6, as a function of GCA, with each sample divided into quintiles, from lowest to highest GCA. The plots are shown in Fig. 7. While GCA level was weakly, and in Lifebrain even inversely related to atrophy in these regions in the younger group, the expected trajectories for the older group were relatively consistently ordered so that persons with a higher GCA level had less decline, especially of cortical thickness. The GCA quintile differences are more pronounced in the older group, suggesting the latter half of the lifespan is driving the interaction. As one outlier in the older group in Lifebrain was noted as having a high cortical thickness value for age at the first timepoint in the region of interest, we carefully checked this segmentation, but found no sign of flawed segmentation, and thus decided to keep this person in analyses.