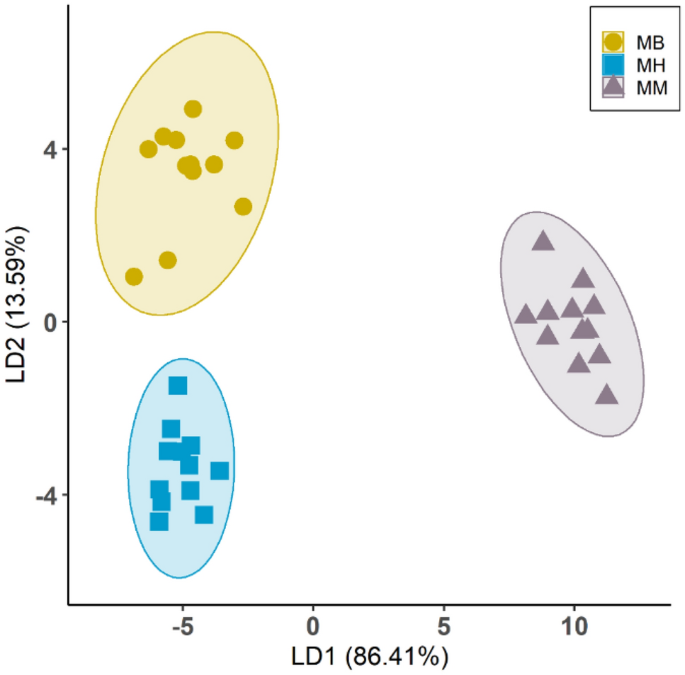
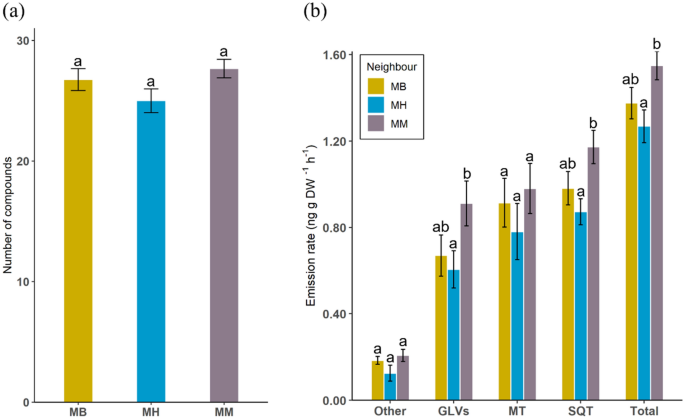
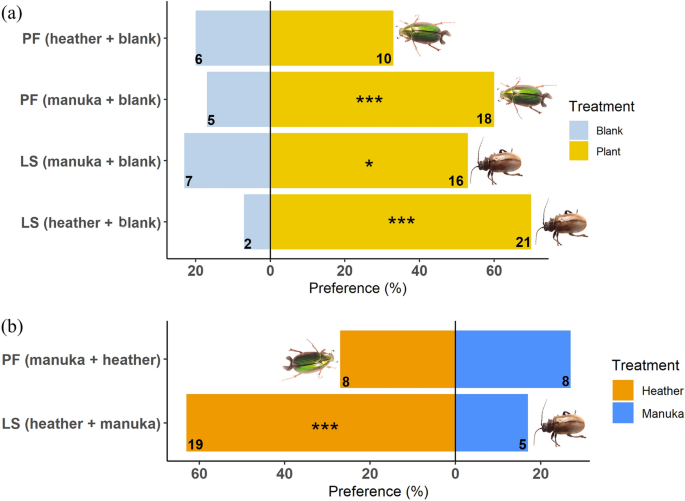
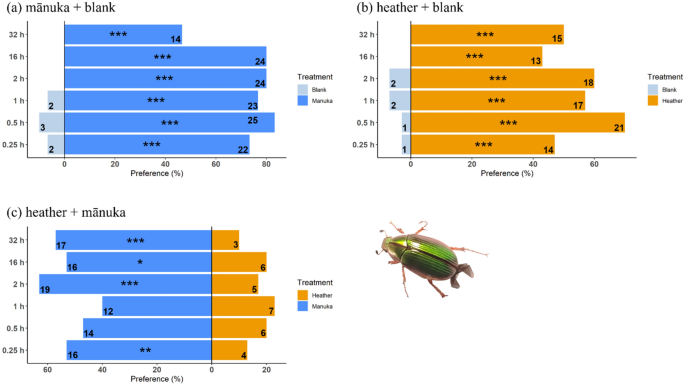
-
Mack, R. N. et al. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 10, 689–710 (2000).
Google Scholar
-
Turbelin, A. J., Malamud, B. D. & Francis, R. A. Mapping the global state of invasive alien species: Patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 26, 78–92 (2017).
Google Scholar
-
Jackson, M. C. Interactions among multiple invasive animals. Ecology 96, 2035–2041 (2015).
CAS
PubMed
Google Scholar
-
Rodriguez, L. F. Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biol. Invasions 8, 927–939 (2006).
Google Scholar
-
Duenas, M. A. et al. The role played by invasive species in interactions with endangered and threatened species in the United States: A systematic review. Biodivers. Conserv. 27, 3171–3183 (2018).
Google Scholar
-
Weidenhamer, J. D. & Callaway, R. M. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J. Chem. Ecol. 36, 59–69 (2010).
CAS
PubMed
Google Scholar
-
Bajwa, A. A., Chauhan, B. S., Farooq, M., Shabbir, A. & Adkins, S. W. What do we really know about alien plant invasion? A review of the invasion mechanism of one of the world’s worst weeds. Planta 244, 39–57 (2016).
CAS
PubMed
Google Scholar
-
Tallamy, D. W., Narango, D. L. & Mitchell, A. B. Do non-native plants contribute to insect declines?. Ecol. Entomol. 46, 729–742. https://doi.org/10.1111/een.12973 (2021).
Article
Google Scholar
-
Bezemer, T. M., Harvey, J. A. & Cronin, J. T. Response of native insect communities to invasive plants. Annu. Rev. Entomol. 59, 119 (2014).
CAS
PubMed
Google Scholar
-
Cheng, F. & Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 6, 1020 (2015).
PubMed
PubMed Central
Google Scholar
-
Kalisz, S., Kivlin, S. N. & Bialic-Murphy, L. Allelopathy is pervasive in invasive plants. Biol. Invasions 23, 367–371 (2021).
Google Scholar
-
Pyšek, P. et al. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Change Biol. 18, 1725–1737 (2012).
ADS
Google Scholar
-
Zhang, P., Li, B., Wu, J. & Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 22, 200–210 (2019).
ADS
PubMed
Google Scholar
-
Dudareva, N., Klempien, A., Muhlemann, J. K. & Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 198, 16–32 (2013).
CAS
PubMed
Google Scholar
-
Clavijo McCormick, A. Can plant–natural enemy communication withstand disruption by biotic and abiotic factors?. Ecol. Evol. 6, 8569–8582 (2016).
PubMed
PubMed Central
Google Scholar
-
Bruce, T. J., Wadhams, L. J. & Woodcock, C. M. Insect host location: A volatile situation. Trends Plant Sci. 10, 269–274 (2005).
CAS
PubMed
Google Scholar
-
Clavijo McCormick, A., Unsicker, S. B. & Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 17, 303–310 (2012).
CAS
PubMed
Google Scholar
-
Baldwin, I. T., Halitschke, R., Paschold, A., Von Dahl, C. C. & Preston, C. A. Volatile signaling in plant–plant interactions: “Talking trees” in the genomics era. Science 311, 812–815 (2006).
ADS
CAS
PubMed
Google Scholar
-
Kegge, W. & Pierik, R. Biogenic volatile organic compounds and plant competition. Trends Plant Sci. 15, 126–132 (2010).
CAS
PubMed
Google Scholar
-
Effah, E., Holopainen, J. K. & Clavijo McCormick, A. Potential roles of volatile organic compounds in plant competition. Perspect. Plant Ecol. Evol. Syst. 38, 58–63 (2019).
Google Scholar
-
Kigathi, R. N., Weisser, W. W., Reichelt, M., Gershenzon, J. & Unsicker, S. B. Plant volatile emission depends on the species composition of the neighboring plant community. BMC Plant Biol. 19, 1–17 (2019).
Google Scholar
-
Karban, R., Wetzel, W. C., Shiojiri, K., Pezzola, E. & Blande, J. D. Geographic dialects in volatile communication between sagebrush individuals. Ecology 97, 2917–2924 (2016).
PubMed
Google Scholar
-
Wheeler, G. S., David, A. S. & Lake, E. C. Volatile chemistry, not phylogeny, predicts host range of a biological control agent of Old-World climbing fern. Biol. Control 159, 104636 (2021).
CAS
Google Scholar
-
Li, N. et al. Manipulating two olfactory cues causes a biological control beetle to shift to non-target plant species. J. Ecol. 105, 1534–1546 (2017).
CAS
Google Scholar
-
Buddenhagen, C. E. Broom Control Monitoring at Tongariro National Park (Department of Conservation Wellington, 2000).
Google Scholar
-
Hayes, L. et al. Biocontrol of Weeds: Achievements to Date and Future Outlook. Ecosystem services in New Zealand-conditions and trends Vol. 2, 375–385 (Manaaki Whenua Press, 2013).
Google Scholar
-
Bagnall, A. Heather at Tongariro. A study of a weed introduction. Tussock Grasslands Mt. Lands Inst. Rev 41, 17–21 (1982).
Google Scholar
-
Chapman, H. M. & Bannister, P. The spread of heather, Calluna vulgaris (L.) Hull, into indigenous plant communities of Tongariro National Park. N. Z. J. Ecol. 7–16 (1990).
-
Effah, E. et al. Effects of two invasive weeds on arthropod community structure on the Central Plateau of New Zealand. Plants 9, 919 (2020).
CAS
PubMed Central
Google Scholar
-
Keesing, V. F. Impacts of invasion on community structure: habitat and invertebrate assemblage responses to Calluna vulgaris (L.) Hull invasion, in Tongariro National Park, New Zealand, Massey University Palmerston North, New Zealand, (1995).
-
Peterson, P. G., Fowler, S. V. & Barrett, P. Is the poor establishment and performance of heather beetle in Tongariro National Park due to the impact of parasitoids predators or disease. N. Z. Plant Prot. 57, 89–93. https://doi.org/10.30843/nzpp.2004.57.6977 (2004).
Article
Google Scholar
-
Ajpark. The brands and the bees: trade marks and the mānuka challenge for honey businesses, https://www.ajpark.com/insights/the-brands-and-the-bees-trade-marks-and-the-manuka-challenge-for-honey-businesses/#:~:text=M%C4%81nuka%20is%20a%20taonga%20species,may%20be%20offensive%20to%20M%C4%81ori (2021).
-
Effah, E. et al. Seasonal and environmental variation in volatile emissions of the New Zealand native plant Leptospermum scoparium in weed-invaded and non-invaded sites. Sci. Rep. 10, 1–11 (2020).
Google Scholar
-
Effah, E., Min Tun, K., Rangiwananga, N. & Clavijo McCormick, A. Mānuka clones differ in their volatile profiles: Potential implications for plant defence, pollinator attraction and bee products. Agronomy 12, 169 (2022).
CAS
Google Scholar
-
Effah, E. et al. Natural variation in volatile emissions of the invasive weed Calluna vulgaris in New Zealand. Plants 9, 283 (2020).
CAS
PubMed Central
Google Scholar
-
Team, R. C. R: A language and environment for statistical computing (R Foundation for Statistical Computing, 2021).
-
Ripley, B. et al. Package ‘mass’. Cran r 538, 113–120 (2013).
Google Scholar
-
Chen, B. M., Liao, H. X., Chen, W. B., Wei, H. J. & Peng, S. L. Role of allelopathy in plant invasion and control of invasive plants. Allelopathy J 41, 155–166 (2017).
Google Scholar
-
Ninkovic, V., Markovic, D. & Rensing, M. Plant volatiles as cues and signals in plant communication. Plant Cell Environ. 44, 1030–1043 (2021).
CAS
PubMed
Google Scholar
-
Holopainen, J. K. Multiple functions of inducible plant volatiles. Trends Plant Sci. 9, 529–533 (2004).
CAS
PubMed
Google Scholar
-
Rhoades, D. F. Responses of alder and willow to attack by tent caterpillars and webworms: evidence for pheromonal sensitivity of willows. In Plant Resistance to Insects (ed. Hedin, P. A.) 55–68 (American Chemical Society, 1983).
Google Scholar
-
Hedin, P. A. Plant Resistance to Insects (American Chemical Society, 1983).
Google Scholar
-
Heil, M. & Karban, R. Explaining evolution of plant communication by airborne signals. Trends Ecol. Evol. 25, 137–144 (2010).
PubMed
Google Scholar
-
Barbosa, P. et al. Associational resistance and associational susceptibility: Having right or wrong neighbors. Annu. Rev. Ecol. Evol. Syst. 40, 1 (2009).
Google Scholar
-
Kigathi, R. N., Weisser, W. W., Veit, D., Gershenzon, J. & Unsicker, S. B. Plants suppress their emission of volatiles when growing with conspecifics. J. Chem. Ecol. 39, 537–545 (2013).
CAS
PubMed
Google Scholar
-
Peñuelas, J. & Llusià, J. Influence of intra-and inter-specific interference on terpene emission by Pinus halepensis and Quercus ilex seedlings. Biol. Plant. 41, 139–143 (1998).
Google Scholar
-
Ormeno, E., Fernandez, C. & Mévy, J.-P. Plant coexistence alters terpene emission and content of Mediterranean species. Phytochemistry 68, 840–852 (2007).
CAS
PubMed
Google Scholar
-
Himanen, S. J. et al. Birch (Betula spp.) leaves adsorb and re-release volatiles specific to neighbouring plants—A mechanism for associational herbivore resistance?. New Phytol. 186, 722–732 (2010).
CAS
PubMed
Google Scholar
-
Kessler, A. & Kalske, A. Plant secondary metabolite diversity and species interactions. Annu. Rev. Ecol. Evol. Syst. 49, 115–138 (2018).
Google Scholar
-
Quintana-Rodriguez, E. et al. Plant volatiles cause direct, induced and associational resistance in common bean to the fungal pathogen Colletotrichum lindemuthianum. J. Ecol. 103, 250–260 (2015).
CAS
Google Scholar
-
Loreto, F. & D’Auria, S. How do plants sense volatiles sent by other plants? Trends Plant Sci. (2021).
-
Giordano, D., Facchiano, A., D’Auria, S. & Loreto, F. A hypothesis on the capacity of plant odorant-binding proteins to bind volatile isoprenoids based on in silico evidences. Elife 10, e66741 (2021).
CAS
PubMed
PubMed Central
Google Scholar
-
Ninkovic, V., Markovic, D. & Dahlin, I. Decoding neighbour volatiles in preparation for future competition and implications for tritrophic interactions. Perspect. Plant Ecol. Evol. Syst. 23, 11–17 (2016).
Google Scholar
-
Kegge, W. et al. Red: far-red light conditions affect the emission of volatile organic compounds from barley (Hordeum vulgare), leading to altered biomass allocation in neighbouring plants. Ann. Bot. 115, 961–970 (2015).
CAS
PubMed
PubMed Central
Google Scholar
-
Gershenzon, J. Metabolic costs of terpenoid accumulation in higher plants. J. Chem. Ecol. 20, 1281–1328 (1994).
CAS
PubMed
Google Scholar
-
Anderson, P., Sadek, M., Larsson, M., Hansson, B. & Thöming, G. Larval host plant experience modulates both mate finding and oviposition choice in a moth. Anim. Behav. 85, 1169–1175 (2013).
Google Scholar
-
Cunningham, J. P., Moore, C. J., Zalucki, M. P. & West, S. A. Learning, odour preference and flower foraging in moths. J. Exp. Biol. 207, 87–94 (2004).
PubMed
Google Scholar
-
McCormick, A. C., Reinecke, A., Gershenzon, J. & Unsicker, S. B. Feeding experience affects the behavioral response of polyphagous gypsy moth caterpillars to herbivore-induced poplar volatiles. J. Chem. Ecol. 42, 382–393 (2016).
CAS
PubMed
PubMed Central
Google Scholar
-
Proffit, M., Khallaf, M. A., Carrasco, D., Larsson, M. C. & Anderson, P. ‘Do you remember the first time?’ Host plant preference in a moth is modulated by experiences during larval feeding and adult mating. Ecol. Lett. 18, 365–374 (2015).
PubMed
Google Scholar
-
Mayhew, P. J. Herbivore host choice and optimal bad motherhood. Trends Ecol. Evol. 16, 165–167 (2001).
PubMed
Google Scholar
-
Jackson, T. et al. Anticipating the unexpected–managing pasture pest outbreaks after large-scale land conversion (New Zealand Grassland Association, 2012).
-
Townsend, R. J., Dunbar, J. E. & Jackson, T. A. Flight behaviour of the manuka chafers, Pyronota festiva (Fabricius) and Pyronota setosa (Given) (Coleoptera: Melolonthinae), on the flipped soils of Cape Foulwind on the West Coast of New Zealand. N. Z. Plant Prot. 71, 255–261. https://doi.org/10.30843/nzpp.2018.71.175 (2018).
Article
Google Scholar
-
Ferguson, C. M. et al. Quantifying the economic cost of invertebrate pests to New Zealand’s pastoral industry. N. Z. J. Agric. Res. 62, 255–315 (2019).
Google Scholar
-
Cunningham, J. Can mechanism help explain insect host choice?. J. Evol. Biol. 25, 244–251 (2012).
CAS
PubMed
Google Scholar
-
Syrett, P., Smith, L. A., Bourner, T. C., Fowler, S. V. & Wilcox, A. A European pest to control a New Zealand weed: Investigating the safety of heather beetle, Lochmaea suturalis (Coleoptera: Chrysomelidae) for biological control of heather, Calluna vulgaris. Bull. Entomol. Res. 90, 169–178. https://doi.org/10.1017/S0007485300000286 (2000).
CAS
Article
PubMed
Google Scholar
-
Fowler, S., Harman, H., Memmott, J., Peterson, P. & Smith, L. In Proceedings of the XII International Symposium on Biological Control of Weeds (eds Julien, M. H. et al.) 495–502.
-
Fowler, S. V. et al. Investigating the poor performance of heather beetle, Lochmaea suturalis (Thompson) (Coleoptera: Chrysomelidae), as a weed biocontrol agent in New Zealand: Has genetic bottlenecking resulted in small body size and poor winter survival?. Biol. Control 87, 32–38 (2015).
Google Scholar
-
Effah, E. et al. Herbivory and attenuated UV radiation affect volatile emissions of the invasive weed Calluna vulgaris. Molecules 25, 3200 (2020).
CAS
PubMed Central
Google Scholar
-
Pearson, D. E. & Callaway, R. M. Indirect nontarget effects of host-specific biological control agents: Implications for biological control. Biol. Control 35, 288–298 (2005).
Google Scholar
-
Rand, T. A. & Louda, S. M. Exotic weed invasion increases the susceptibility of native plants to attack by a biocontrol herbivore. Ecology 85, 1548–1554. https://doi.org/10.1890/03-3067 (2004).
Article
Google Scholar