







Across the U.S., nighttime lights start getting brighter on “Black Friday,” the day after Thanksgiving, and remain through New Year’s Day.
In the Middle East, night lights shine 50 per cent brighter during the holy month of Ramadan.
These observations were made by NASA scientists in 2014, who used data collected by the Suomi NPP satellite to analyze nighttime light activity over 70 U.S. cities.
The purpose of the study, led by Miguel Román of NASA’s Goddard Space Flight Center, was to determine patterns in urban energy use, which is a key component of greenhouse gas emissions.
They discovered light intensity increased by 30 to 50 per cent in the suburbs and outskirts of major cities during the holiday season. Lights in the central urban areas also increase during the holidays, but not as much.
“More than 70 percent of greenhouse gas emissions come from urban areas,” Román said in a 2014 statement.

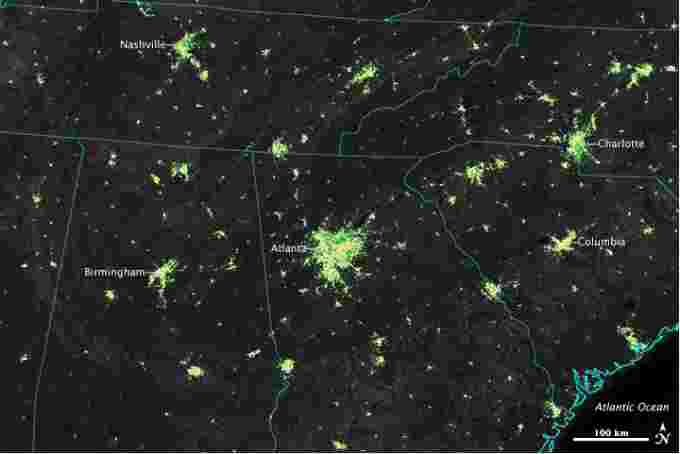
Holiday season lighting increases seen over parts of the U.S. between 2012 and 2014. Courtesy: NASA.
“If we’re going to reduce these emissions, then we’ll have to do more than just use energy-efficient cars and appliances. We also need to understand how dominant social phenomena, the changing demographics of urban centers, and socio-cultural settings affect energy-use decisions.”
CELEBRATING CHRISTMAS THE ENVIRONMENTALLY-FRIENDLY WAY
Canadians are becoming increasingly aware of the environmental impact of the holidays.
There are several ways holiday lights can be an ecological drain. In addition to contributing to greenhouse gas emissions, the devices contribute to light pollution which can be disruptive to nocturnal wildlife.
While most holiday lights are recyclable, countless strands end up in the landfill alongside the estimated 540,000 tonnes of gift packaging, 2.6 billion gift cards, and 6 million rolls of tape Canadians toss out each holiday season.
ECO-FRIENDLY GIFT GIVING
Check out the video below for some tips on how to be a little greener with your gifts this year.









