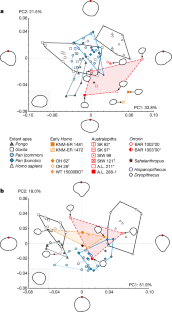
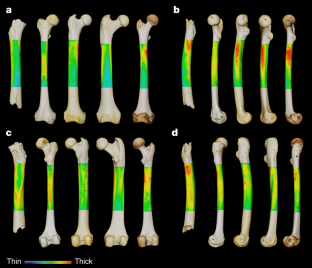
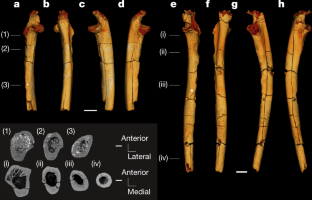
-
Senut, B. et al. First hominid from the Miocene (Lukeino formation, Kenya). C. R. Acad. Sci. Paris IIA 332, 137–144 (2001).
Article
Google Scholar
-
Pickford, M., Senut, B., Gommery, D. & Treil, J. Bipedalism in Orrorin tugenensis revealed by its femora. C. R. Palevol. 1, 191–203 (2002).
Article
Google Scholar
-
Almécija, S. et al. The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nat. Commun. 4, 2888 (2013).
ADS
PubMed
Article
CAS
Google Scholar
-
Richmond, B. G. & Jungers, W. L. Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism. Science 319, 1662–1665 (2008).
ADS
CAS
PubMed
Article
Google Scholar
-
Brunet, M. et al. A new hominid from the Upper Miocene of Chad, Central Africa. Nature 418, 145–151 (2002).
ADS
CAS
PubMed
Article
Google Scholar
-
Zollikofer, C. P. et al. Virtual cranial reconstruction of Sahelanthropus tchadensis. Nature 434, 755–759 (2005).
ADS
CAS
PubMed
Article
Google Scholar
-
Lebatard, A. E. et al. Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene hominids from Chad. Proc. Natl Acad. Sci. USA 105, 3226–3231 (2008).
ADS
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Brunet, M. et al. New material of the earliest hominid from the Upper Miocene of Chad. Nature 434, 752–755 (2005).
ADS
CAS
PubMed
Article
Google Scholar
-
Vignaud, P. et al. Geology and palaeontology of the Upper Miocene Toros-Menalla hominid locality, Chad. Nature 418, 152–155 (2002).
ADS
CAS
PubMed
Article
Google Scholar
-
Heaton, J. L. et al. The long limb bones of the StW 573 Australopithecus skeleton from Sterkfontein Member 2: descriptions and proportions. J. Hum. Evol. 133, 167–197 (2019).
PubMed
Article
Google Scholar
-
Ruff, C. B. Biomechanics of the hip and birth in early Homo. Am. J. Phys. Anthrop. 98, 527–574 (1995).
CAS
PubMed
Article
Google Scholar
-
Böhme, M. et al. A new Miocene ape and locomotion in the ancestor of great apes and humans. Nature 575, 489–493 (2019).
ADS
PubMed
Article
CAS
Google Scholar
-
Williams, S. A. et al. Reevaluating bipedalism in Danuvius. Nature 586, E1–E3 (2020).
ADS
Article
CAS
Google Scholar
-
Pina, M. Unravelling the Positional Behaviour of Fossil Hominoids Morphofunctional and Structural Analysis of the Primate Hindlimb. PhD thesis, Universitat Autònoma de Barcelona (2016).
-
Lovejoy, C. O., Suwa, G., Spurlock, L., Asfaw, B. & White, T. D. The pelvis and femur of Ardipithecus ramidus: the emergence of upright walking. Science 326, 71e1–71e6 (2009).
PubMed
Google Scholar
-
Prost, J. H. A definitional system for the classification of primate locomotion. Am. Anthropol. 67, 1198–1214 (1965).
Article
Google Scholar
-
Puymerail, L. The functionally-related signatures characterizing the endostructural organisation of the femoral shaft in modern humans and chimpanzee. C. R. Palevol. 12, 223–231 (2013).
Article
Google Scholar
-
Galik, K. et al. External and internal morphology of the BAR 1002′00 Orrorin tugenensis femur. Science 305, 1450–1453 (2004).
ADS
CAS
PubMed
Article
Google Scholar
-
Ohman, J. C., Lovejoy, C. O. & White, T. D. Questions about Orrorin femur. Science 307, 845 (2005).
CAS
PubMed
Article
Google Scholar
-
Puymerail, L. Caractérisation de l’endostructure et des propriétés biomécaniques de la diaphyse fémorale: la signature de la bipédie et la reconstruction des paléo-répertoires posturaux et locomoteurs des hominines (Paris, Muséum National d’Histoire Naturelle, 2011).
-
Wallace, I. J. et al. Functional significance of genetic variation underlying limb bone diaphyseal structure. Am. J. Phys. Anthropol. 143, 21–30 (2010).
PubMed
PubMed Central
Article
Google Scholar
-
Lieberman, D. E., Polk, J. D. & Demes, B. Predicting long bone loading from cross‐sectional geometry. Am. J. Phys. Anthropol. 123, 156–171 (2004).
PubMed
Article
Google Scholar
-
Grabowski, M., Hatala, K. G., Jungers, W. L. & Richmond, B. G. Body mass estimates of hominin fossils and the evolution of human body size. J. Hum. Evol. 85, 75–93 (2015).
PubMed
Article
Google Scholar
-
Ruff, C. B., Burgess, M. L., Squyres, N., Junno, J. A. & Trinkaus, E. Lower limb articular scaling and body mass estimation in Pliocene and Pleistocene hominins. J. Hum. Evol. 115, 85–111 (2018).
PubMed
Article
Google Scholar
-
Nadell, J. Ontogeny and Adaptation: A Cross-Sectional Study of Primate Limb Elements. PhD thesis, Durham University (2017).
-
Nakatsukasa, M., Pickford, M., Egi, N. & Senut, B. Femur length, body mass, and stature estimates of Orrorin tugenensis, a 6 Ma hominid from Kenya. Primates 48, 171–178 (2007).
PubMed
Article
Google Scholar
-
Lovejoy, C. O. et al. The great divides: Ardipithecus ramidus reveals the postcrania of our last common ancestors with African apes. Science 326, 73–106 (2009).
ADS
Article
CAS
Google Scholar
-
Jungers, W. L., Grabowski, M., Hatala, K. G. & Richmond, B. G. The evolution of body size and shape in the human career. Phil. Trans. R. Soc. B 371, 20150247 (2016).
PubMed
PubMed Central
Article
Google Scholar
-
Kuperavage, A., Pokrajac, D., Chavanaves, S. & Eckhardt, R. B. Earliest known hominin calcar femorale in Orrorin tugenensis provides further internal anatomical evidence for origin of human bipedal locomotion. Anat. Rec. 301, 1834–1839 (2018).
CAS
Article
Google Scholar
-
Clark, J. D. et al. Palaeoanthropological discoveries in the middle Awash Valley, Ethiopia. Nature 307, 423–428 (1984).
ADS
Article
Google Scholar
-
Hammer, A. The calcar femorale: a new perspective. J. Orthop. Surg. 27, 2309499019848778 (2019).
Article
Google Scholar
-
Zhang, Q. et al. The role of the calcar femorale in stress distribution in the proximal femur. Orthop. Surg. 1, 311–316 (2009).
PubMed
PubMed Central
Article
Google Scholar
-
Haile-Selassie, Y., Suwa, G. & White, T. Hominidae. in Ardipithecus kadabba: Late Miocene Evidence From The Middle Awash, Ethiopia (eds Haile-Selassie, Y. & WoldeGabriel, G.) 159–236 (Univ. California Press, 2009).
-
Araiza, I., Meyer, M. R. & Williams, S. A. Is ulna curvature in the StW 573 (‘Little Foot’) Australopithecus natural or pathological? J. Hum. Evol. 151, 102927 (2021).
PubMed
Article
Google Scholar
-
Drapeau, M. S. M., Ward, C. V., Kimbel, W. H., Johanson, D. C. & Rak, Y. Associated cranial and forelimb remains attributed to Australopithecus afarensis from Hadar, Ethiopia. J. Hum. Evol. 48, 593–642 (2005).
CAS
PubMed
Article
Google Scholar
-
Henderson, K., Pantinople, J., McCabe, K., Richards, H. L. & Milne, N. Forelimb bone curvature in terrestrial and arboreal mammals. PeerJ 5, e3229 (2017).
PubMed
PubMed Central
Article
Google Scholar
-
Drapeau, M. S. M. Functional anatomy of the olecranon process in hominoids and Plio-Pleistocene hominins. Am. J. Phys. Anthropol. 124, 297–314 (2004).
PubMed
Article
Google Scholar
-
Milne, N. & Granatosky, M. C. Ulna curvature in arboreal and terrestrial primates. J. Mammal. Evol. 28, 897–909 (2021).
Article
Google Scholar
-
Carlson, K. J. et al. Role of nonbehavioral factors in adjusting long bone diaphyseal structure in free-ranging Pan troglodytes. Int. J. Primatol. 29, 1401–1420 (2008).
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Schmitt, D. Mediolateral reaction forces and forelimb anatomy in quadrupedal primates: implications for interpreting locomotor behavior in fossil primates. J. Hum. Evol. 44, 47–58 (2003).
PubMed
Article
Google Scholar
-
Cartmill, M. & Milton, K. The iorisiform wrist joint and the evolution of “brachiating” adaptations in the Hominoidea. Am. J. Phys. Anthrop. 47, 249–272 (1977).
CAS
PubMed
Article
Google Scholar
-
Hunt, K. D. et al. Standardized descriptions of primate locomotor and postural modes. Primates 37, 363–387 (1996).
Article
Google Scholar
-
Sarmiento, E. E. Anatomy of the hominoid wrist joint: its evolutionary and functional implications. Int. J. Primatol. 9, 281–345 (1988).
Article
Google Scholar
-
Begun, D. R. Phyletic diversity and locomotion in primitive European hominids. Am. J. Phys. Anthropol. 87, 311–340 (1992).
CAS
PubMed
Article
Google Scholar
-
Lovejoy, C. O., Simpson, S. W., White, T. D., Asfaw, B. & Suwa, G. Careful climbing in the Miocene: the forelimbs of Ardipithecus ramidus and humans are primitive. Science 326, 70e1–70e8 (2009).
PubMed
Google Scholar
-
Drapeau, M. S. M. Articular morphology of the proximal ulna in extant and fossil hominoids and hominins. J. Hum. Evol. 55, 86–102 (2008).
PubMed
Article
Google Scholar
-
Alba, D. M., Almécija, S., Casanovas-Vilar, I., Méndez, J. M. & Moyà-Solà, S. A partial skeleton of the fossil great ape Hispanopithecus laietanus from Can Feu and the mosaic evolution of crown-hominoid positional behaviors. PLoS ONE 7, e39617 (2012).
ADS
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Tuttle, R. H. Knuckle‐walking and the evolution of hominoid hands. Am. J. Phys. Anthrop. 26, 171–206 (1967).
Article
Google Scholar
-
Crompton, R. H., Vereecke, E. E. & Thorpe, S. K. Locomotion and posture from the common hominoid ancestor to fully modern hominins, with special reference to the last common panin/hominin ancestor. J. Anat. 212, 501–543 (2008).
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Stern, J. T. & Susman, R. L. in Origine(s) de la Bipédie chez les Hominidés (eds Coppens, Y. & Senut, B.) 99–111 (CNRS, 1991).
-
Kozma, E. E. et al. Hip extensor mechanics and the evolution of walking and climbing capabilities in humans, apes, and fossil hominins. Proc. Natl Acad. Sci. USA 115, 4134–4139 (2018).
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Macchiarelli, R., Bergeret-Medina, A., Marchi, D. & Wood, B. Nature and relationships of Sahelanthropus tchadensis. J. Hum. Evol. 149, 102898 (2020).
PubMed
Article
Google Scholar
-
Gommery, D. & Senut, B. The terminal thumb phalanx of Orrorin tugenensis (Upper Miocene of Kenya). Geobios 39, 372–384 (2006).
Article
Google Scholar
-
Kirscher, U. et al. Age constraints for the Trachilos footprints from Crete. Sci. Rep. 11, 19427 (2021).
ADS
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Meldrum, J. & Sarmiento, E. Comments on possible Miocene hominin footprints. Proc. Geol. Assoc. 129, 577–580 (2018).
Article
Google Scholar
-
Guy, F. et al. Morphological affinities of the Sahelanthropus tchadensis (late Miocene hominid from Chad) cranium. Proc. Natl Acad. Sci. USA 102, 18836–18841 (2005).
ADS
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Neaux, D. et al. Relationship between foramen magnum position and locomotion in extant and extinct hominoids. J. Jum. Evol. 113, 1–9 (2017).
Google Scholar
-
Pilbeam, D. R. & Lieberman, D. E. in Chimpanzees and Human Evolution (eds Muller, M. N., Wrangham, R. W. & Pilbeam, D. R.) 22–141 (Belknap Harvard, 2017).
-
Senut, B., Pickford, M., Gommery, D. & Ségalen, L. Palaeoenvironments and the origin of hominid bipedalism. Hist. Biol. 30, 284–296 (2018).
Article
Google Scholar
-
WoldeGabriel, G. et al. Geology and palaeontology of the late Miocene Middle Awash valley, Afar rift, Ethiopia. Nature 412, 175–178 (2001).
ADS
CAS
PubMed
Article
Google Scholar
-
White, T. D. et al. Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science 326, 67–93 (2009).
ADS
Article
CAS
Google Scholar
-
Barboni, D., Ashley, G. M., Bourel, B., Arraiz, H. & Mazur, J. C. Springs, palm groves, and the record of early hominins in Africa. Rev. Palaeobot. Palynol. 266, 23–41 (2019).
Article
Google Scholar
-
Novello, A. et al. Phytoliths indicate significant arboreal cover at Sahelanthropus type locality TM266 in northern Chad and a decrease in later sites. J. Hum. Evol. 106, 66–83 (2017).
PubMed
Article
Google Scholar
-
Steiper, M. E. & Seiffert, E. R. Evidence for a convergent slowdown in primate molecular rates and its implications for the timing of early primate evolution. Proc. Natl Acad. Sci. USA 109, 6006–6011 (2012).
ADS
CAS
PubMed
PubMed Central
Article
Google Scholar
-
Püschel, H. P., Bertrand, O. C., O’reilly, J. E., Bobe, R. & Püschel, T. A. Divergence-time estimates for hominins provide insight into encephalization and body mass trends in human evolution. Nat. Ecol. Evol. 5, 808–819 (2021).
PubMed
Article
Google Scholar
-
Besenbacher, S., Hvilsom, C., Marques-Bonet, T., Mailund, T. & Schierup, M. H. Direct estimation of mutations in great apes reconciles phylogenetic dating. Nat. Ecol. Evol. 3, 286–292 (2019).
PubMed
Article
Google Scholar
-
McBrearty, S. & Jablonski, N. G. First fossil chimpanzee. Nature 437, 105–108 (2005).
ADS
CAS
PubMed
Article
Google Scholar
-
DeSilva, J., Shoreman, E. & MacLatchy, L. A fossil hominoid proximal femur from Kikorongo Crater, southwestern Uganda. J. Hum. Evol. 50, 687–695 (2006).
PubMed
Article
Google Scholar
-
Ishida, H. & Pickford, M. A new late Miocene hominoid from Kenya: Samburupithecus kiptalami gen. et sp. nov. C. R. Acad. Sci. Paris 325, 823–829 (1997).
Google Scholar
-
Kunimatsu, Y. et al. A new late Miocene great ape from Kenya and its implications for the origins of African great apes and humans. Proc. Natl Acad. Sci. USA 104, 19220–19225 (2007).
ADS
PubMed
PubMed Central
Article
Google Scholar
-
Suwa, G., Kono, R. T., Katoh, S., Asfaw, B. & Beyene, Y. A new species of great ape from the late Miocene epoch in Ethiopia. Nature 448, 921–924 (2007).
ADS
CAS
PubMed
Article
Google Scholar
-
Katoh, S. et al. New geological and palaeontological age constraint for the gorilla–human lineage split. Nature 530, 215–218 (2016).
ADS
CAS
PubMed
Article
Google Scholar
-
Begun, D. R. in Handbook of Paleoanthropology (eds Henke, W. & Tattersall, I.) 1261–1332 (Springer-Verlag, 2015).
-
Mongle, C. S., Strait, D. S. & Grine, F. E. Expanded character sampling underscores phylogenetic stability of Ardipithecus ramidus as a basal hominin. J. Hum. Evol. 131, 28–39 (2019).
PubMed
Article
Google Scholar
-
Wood, B. & Harrison, T. The evolutionary context of the first hominins. Nature 470, 347–352 (2011).
ADS
CAS
PubMed
Article
Google Scholar
-
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
CAS
PubMed
Article
Google Scholar
-
Mongle, C. S., Wallace, I. J. & Grine, F. E. Cross-sectional structural variation relative to midshaft along hominine diaphyses. II. The hind limb. Am. J. Phys. Anthropol. 158, 398–407 (2015).
PubMed
Article
Google Scholar
-
Puymerail, L. et al. Structural analysis of the Kresna 11 Homo erectus femoral shaft (Sangiran, Java). J. Hum. Evol. 63, 741–749 (2012).
PubMed
Article
Google Scholar
-
Ruff, C. B., McHenry, H. M. & Thackeray, J. F. Cross‐sectional morphology of the SK 82 and 97 proximal femora. Am. J. Phys. Anthrop. 109, 509–521 (1999).
CAS
PubMed
Article
Google Scholar
-
Ruff, C. B. Long bone articular and diaphyseal structure in Old World monkeys and apes. I: locomotor effects. Am. J. Phys. Anthrop. 119, 305–342 (2002).
PubMed
Article
Google Scholar
-
Sládek, J. et al. Effect of deriving periosteal and endosteal contours from microCT scans on computation of cross-sectional properties in non-adults: the femur. J. Anat. 233, 381–393 (2018).
PubMed Central
Article
Google Scholar
-
Ruff, C. E., Higgins, R. W., Carlson, K. J. in Hominin Postcranial Remains from Sterkfontein, South Africa, 1936–1995 (eds Zipfel, B., Richmond B. G. & Ward, C.) 307–320 (Oxford Univ. Press, 2020).
-
Rohlf, F. J. tpsDig, digitize landmarks and outlines v.2.05 (Department of Ecology and Evolution, State University of New York at Stony Brook, 2005).
-
Senut, B. Bipédie et climat. C. R. Palevol. 5, 89–98 (2006).
Article
Google Scholar
-
Adams D., Collyer M., Kaliontzopoulou A. & Baken E. geomorph: Geometric Morphometric Analyses of 2D/3D Landmark Data. R package version 4.0. https://cran.r-project.org/package=geomorph (2021).
-
Baken, E. K., Collyer, M. L., Kaliontzopoulou, A. & Adams, D. C. Geomorph v4.0 and gmShiny: enhanced analytics and a new graphical interface for a comprehensive morphometric experience. Meth. Ecol. Evol. 12, 2355–2363 (2021).
Article
Google Scholar
-
The Development Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2021).
-
Bookstein, F. L. Pathologies of between-groups principal components analysis in geometric morphometrics. Evol. Biol. 46, 271–302 (2019).
Article
Google Scholar
-
Cardini, A., O’Higgins, P. & Rohlf, F. J. Seeing distinct groups where there are none: spurious patterns from between-group PCA. Evol. Biol. 46, 303–316 (2019).
Article
Google Scholar