








Two SpaceX Falcon 9 rockets have completed back-to-back Starlink launches less than 24 hours apart, successfully delivering 106 Starlink satellites to low Earth orbit (LEO).
Originally scheduled just a handful of hours apart, slight delays eventually saw Starlink 4-13 and Starlink 4-15 settle on 6:07 pm EDT, May 13th and 4:40 pm EDT, May 14th, respectively. Entering the final stretch, launch preparations went smoothly and both Falcon 9 rockets ultimately lifted off without a hitch.

The series began with Starlink 4-13 on Friday. SpaceX chose Falcon 9 B1063 to support the Starlink launch and the booster did its job well, wrapping up its fifth launch since November 2020 with a rare landing aboard drone ship Of Course I Still Love You (OCISLY). Since SpaceX permanently transferred OCISLY from the East Coast to the West Coast in mid-2021, the drone ship has only supported five booster recoveries. Save for an unusual East Coast Starlink launch in May 2021, Falcon 9 B1061 has also primarily been tasked with supporting SpaceX’s West Coast launch manifest. With only one older pad – Vandenberg Space Force Base’s (VSFB) SLC-4 complex – available to SpaceX, the company’s West Coast Falcon launches are also considerably rarer than its East Coast missions.
SpaceX has also taken to using the pad – which is in an optimal location to launch satellites that orbit Earth’s poles – to launch several batches of Starlink satellites into more ordinary equatorial orbits, essentially augmenting the capabilities of its two Florida launch sites.
Starlink 4-13 and 4-15 were more or less identical, in that regard; both launched 53 Starlink V1.5 satellites into LEO to continue filling out the fourth of five Starlink orbital ‘shells’ that will make up SpaceX’s first licensed constellation. Since SpaceX began Plane 4 (or Group 4) launches in November 2021, the company has now completed 15 missions that carried a total of 860 Starlink V1.5 satellites into orbit. Excluding a solar storm-related fluke that destroyed almost an entire launch worth of satellites, all but 8 remain operational in orbit. According to astronomer Jonathan McDowell’s independent tracking, about 300 Group 4 Starlink satellites have reached operational orbits, while another 500 or so are either raising their orbits or waiting for the right moment to do so.
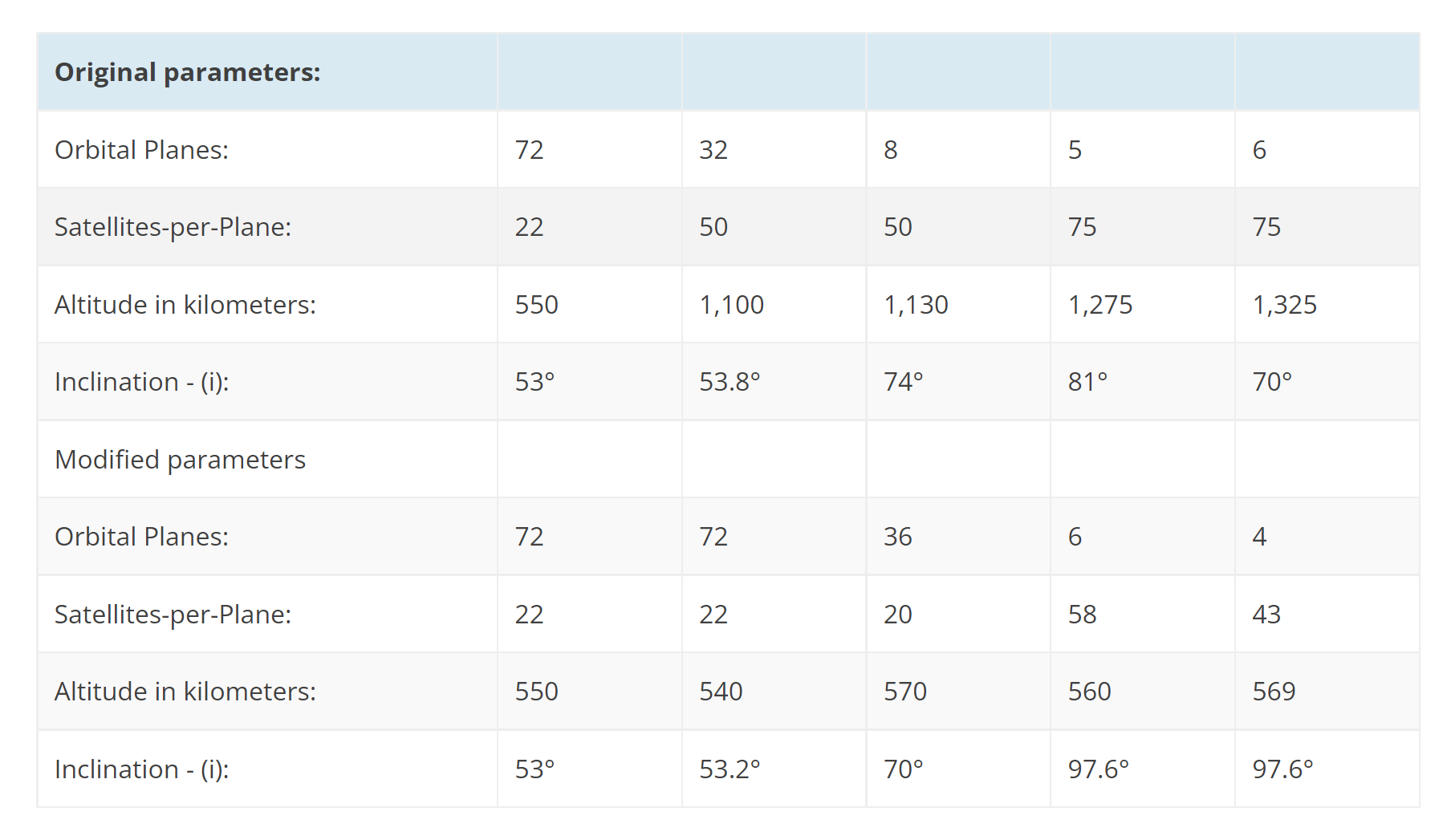
As of May 2022, the first shell or ‘group’ of SpaceX’s first Starlink constellation has about 1500 operational Starlink satellites of a nominal 1584. If all working Group 4 satellites currently in orbit become operational, SpaceX has another ~770 satellites or 15 launches to go to complete the shell (17 to finish Shell 1 and Shell 4). If SpaceX maintains its current six-month launch cadence of one Starlink mission every ~11 days, SpaceX’s first Starlink constellation could have around 3400 working satellites in orbit and be more than three-quarters complete by the end of 2022.
SpaceX, by all appearances, fully intends to push its vehicles and workforce to the absolute limits in 2022 in a bid to complete as many as 60 orbital launches. To launch Starlink 4-15, for example, SpaceX made an unprecedented decision to debut a brand new Falcon 9 booster on the internal mission, demonstrating just how fully its customers have embraced reusability and how much the company wants to expand its fleet of Falcon 9 boosters as quickly as possible.
Following Starlink 4-13 and 4-15, SpaceX has completed 20 launches in the first 19 weeks of 2022 and has another two launches scheduled in the last two weeks of May.

<!–
–>









