








Abstract
The pathogenesis of rheumatoid arthritis (RA) is characterized by a Th17/Treg cell imbalance. A pro-inflammatory cytokine milieu that promotes the continued proliferation of Th17 cells is related to the development of autoinflammation. In RA, T cells have several hallmarks of cellular aging, and they accumulate DNA damage, predisposing to the occurrence of mutations and epigenetic alterations. Since the onset, progression, and treatment response are influenced by a variety of external stressors and environmental factors, this study aimed to evaluate the impact of 8-week yoga practice on disease severity, T cell subsets, markers of T cell ageing and inflammation, epigenetic alterations and gene expression patterns in active RA patients on standard disease-modifying anti-rheumatic drugs (DMARDs). A total of 64 participants with active RA were randomized into 2 groups, yoga group (n = 32) or non-yoga group (n = 32); that were assessed for disease severity, at baseline and after 8 week duration, for Disease Activity Score (DAS28-ESR), T cell subsets [Th17 (CD3+ CD4+ IL17+ RORγt+) cells and Treg (CD3+ CD4+ CD25+ CD127-Foxp3+) cells], markers of T cell aging [aged Th17 cells (CD3+ CD4+ IL17+ RORγt+ CD28−) and aged Treg cells (CD3+ CD4+ CD25+ CD127-Foxp3+ CD28−)], pro-inflammatory markers [IL-6, and IL-17], anti-inflammatory markers [TGF-β, and IL-10], epigenetic alterations [5-methyl cytosine, 5-hydroxymethyl cytosine, and HDAC1] and gene expression patterns [RORγt, FoxP3, IL-17, IL-6, TGF-β, CXCL2, CXCR2, and JUN]. In yoga group, there was a significant improvement in DAS28-ESR scores at the end of 8-weeks of yoga program. The Th17 cells and aged T cell subsets showed a significant decline whereas Treg cell population showed a significant elevation in yoga group. There were significant improvements observed in epigenetic markers as well as inflammatory markers post 8-weeks of yoga practice. The yoga group showed downregulation of RORγt, IL-17, IL-6, CXCL2, CXCR2, and upregulation of FoxP3 and TGF-β transcripts. Yoga enables the maintenance of immune-homeostasis as evident by increased Treg cell population and reduced Th17 cell population. Yoga reduces the rate of immunological aging in T cells, as seen by the reduction in population of aged Th17 cells and aged Treg cells. Yoga positively modifies transcriptome and epigenome by normalization of various inflammatory markers, gene expression patterns and epigenetic alterations. Taken together, yoga reduces RA severity, and aids in immune-modulation and hence can be beneficial as an adjunct therapy.
Introduction
Rheumatoid arthritis (RA) is a chronic inflammatory autoimmune disease of multifactorial origin that develops due to unfavorable coincidence of genetic, immune, and environmental factors1. A fragile state of dynamism persists between pro-inflammatory and anti-inflammatory forces. Loss of tolerance to self-antigens is the hallmark of autoimmune disorders leading to the production of autoantibodies2. The pathogenesis of RA is heavily influenced by immune cells, particularly B-cells, T-cells, and macrophages3. T-cells activate macrophages and fibroblasts to become tissue-destructive cells and release a wide range of cytokines and chemokines to support joint inflammation4.
T helper 17 (Th17) and regulatory T cells (Treg cells) are both differentiated from the same naive CD4+ T cells, but in separate cytokine environments and with distinctive gene expression profiles5. Pro-inflammatory Th17 cells cause joint injury and autoimmune-derived tissue inflammation by inducing proinflammatory cytokines6. Treg cells inhibit the activity of Th17 cells as well as other effector T cells. Treg cells suppress the immune system, maintain self-tolerance, and prevent autoimmune disease by producing anti-inflammatory cytokines7. The retinoic acid-related orphan receptor gamma t (RORγt), a lineage-defining transcription factor unique to Th17 cells that produce interleukin (IL)-17, has been linked to a variety of autoimmune diseases8. Foxp3 (forkhead transcription factor) plays a crucial role as a lineage specification factor of Treg cells that produce transforming growth factor (TGF)-β, which maintains immune tolerance and homeostasis of the immune system9. Th17 cells mediate the pro-inflammatory response through the release of IL-17A and tumor necrosis factor-alpha (TNF-α), which results in tissue destruction as well as damage to bone and cartilage8. In contrast, Treg cells mediate the anti-inflammatory response through the release of IL-10 and TGF-β, which aid in maintenance of immune-tolerance9. The key immune cells, Treg cells are functionally impaired in RA10. The importance of the Th17/Treg cell imbalance in the etiology of RA was highlighted by their opposing behaviors as well as their reciprocal plasticity.
An aberrant Treg cell response with a shift towards a Th17 cell response characterizes the disease onset and course. The relationship between the imbalance of Th17/Treg cells and the production of pro- and anti-inflammatory cytokines is important for the onset and/or course of autoimmunity, chronic inflammation, and articular damage in the joints of RA patients11. Age-inappropriate telomere shortening is a hallmark of premature aging in the T cells of RA patients12. In RA, the rate of immunosenescence is accelerated13. T cells become susceptible to aging as there is enormous proliferative stress due to antigenic exposure, homeostatic proliferation due to thymic involution and their long lifespan as they serve as the carriers of immune memory14. The peculiar features of prematurely aged RA T cells include loss of CD28 expression, shrinking naïve and expanding memory repertoire, shortened telomeres, gain of cytotoxicity, DNA damage accumulation, lack an irreversible cell cycle arrest, more vulnerable to apoptosis, altered tissue trafficking, excess pro-inflammatory cytokine secretion12,15,16. The inefficient DNA damage sensing and repair machinery of RA T cells are linked to premature T cell aging and arthritogenic effector functions16.
Epigenetic alterations like DNA methylation and histone acetylation accumulate with aging and provide a mechanistic link between immunosenescence and development of autoimmune diseases such as RA. T cell landscape is influenced by epigenetic mechanisms such as alteration in the global 5-methyl cytosine (5-mC) DNA, global 5-hydroxymethyl cytosine (5-hmC) DNA and histone deacetylase 1 (HDAC1) levels, which are susceptible to systemic factors, external stressors and environmental stimuli17. Yoga is an integrated ancient mind–body practice that is increasingly recognized to have beneficial effects on immune system functioning18,19,20,21,22,23. Yoga practice may exert positive impact on overall health by boosting cell-mediated and mucosal immunity23. Our previous studies evaluated the role of yoga as an effective intervention to assist the management of RA with respect to various systemic inflammatory biomarkers, immune-modulatory markers, acute phase reactants, clinical symptoms, depression severity, functional status, pain acuity and quality of life19,20,24,25. Yoga aids in regression of inflammatory processes by reducing the pro-inflammatory cytokines and elevating anti-inflammatory cytokines21. Nevertheless, the research on exploration of molecular, cellular and epigenetic aspects following yogic practices is rare, hence a possible mode of action underlying the immune-modulatory effects of yoga needs an active exploration. Further investigation is needed to determine how regular yoga practice impacts the T cell subsets that secrete these cytokines, and the associated transcription factors, gene expression and epigenome. There is currently insufficient data to relate yoga’s multifaceted therapeutic targets and its molecular mode of action with a malfunctioning immune system as in RA.
Keeping in mind the multifactorial etiology, diverse pathogenesis of RA, heterogeneous clinical phenotypes and the therapeutic potential of yoga, we hypothesized that yoga improves clinical outcome in RA by bringing changes in all interconnected biological components and at various levels—molecular, cellular, organ systems, and the person as a whole. With this novel context in mind, this study aimed to investigate the immune-modulatory effects of 8-weeks of yoga practice on disease severity, T cell sub-sets [Th17 (CD3+ CD4+ IL17+ RORγt+) cells and Treg (CD3+ CD4+ CD25+ CD127-Foxp3+) cells], markers of T cell aging [aged Th17 (CD3+ CD4+ IL17+ RORγt+ CD28−) cells and aged Treg (CD3+ CD4+ CD25+ CD127− Foxp3+ CD28−) cells], inflammatory markers [IL-6, IL-17, TGF-β, and IL-10], epigenetic alterations [5-mC, 5-hmC and HDAC1] and gene expression [RORγt, FoxP3, IL-17, IL-6, TGF-β, C‐X‐C motif chemokine ligand 2 (CXCL2), C‐X‐C motif chemokine receptor 2 (CXCR2), and JUN].
Results
Overview of enrollment
A total of 105 individuals were screened for eligibility, out of which 64 were randomized into 2 groups (each group n = 32). Figure 1 shows the CONSORT flowchart of intervention. All 32 randomized participants were included in intent-to-treat analyses of outcome measures.
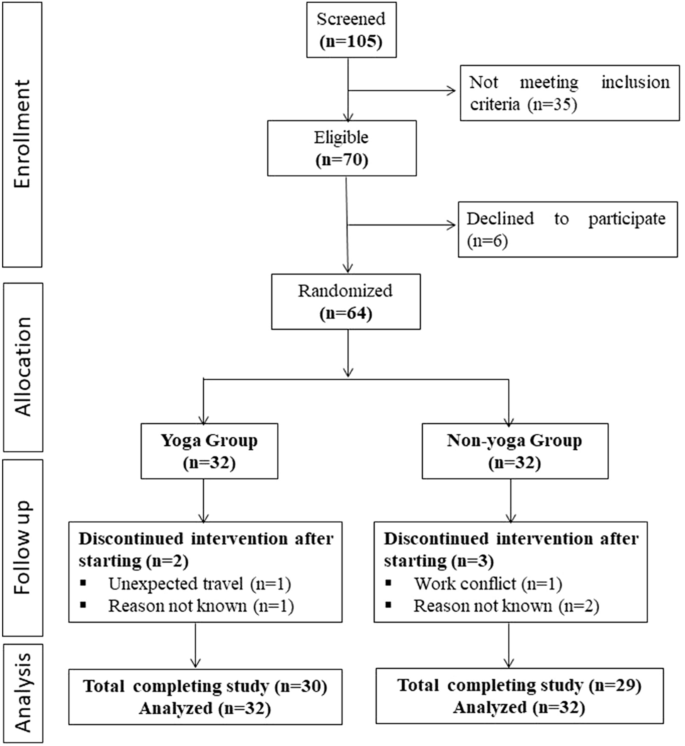
A consort flow diagram of the study.
Participants’ baseline characteristics
Baseline demographic and clinical characteristics of all randomized participants are shown in Table 1. There were no statistically significant differences between the two intervention groups, except stratification of disease severity.
Group × gender interactions
There was no significant difference in mean DAS28-ESR values between males and females at baseline (DAS28-ESR values 4.7 ± 0.9 and 4.6 ± 1.0 mean ± SD, respectively). Separate analyses for males and females were performed to overcome baseline violations in disease activity and to explore further specific gender effects. Interaction effects including group and gender indicated differential responses to yoga for women for DAS28-ESR [F(1,25) = 3.39; p = 0.042], Th17 cells [F(1,25) = 4.06; p = 0.025], Treg cells [F(1,25) = 21.23; p < 0.001], aged Th17 cells [F(1,245 = 14.5; p < 0.001], aged Treg cells [F(1,25) = 13.9; p < 0.001], TGF-β [F(1,25) = 4.62; p = 0.015], 5-mC [F(1,25) = 7.5; p = 0.001] and HDAC1 [F(1,25) = 6.49; p = 0.003]. Clinical improvement was more significant for the women in yoga group [mean between-group difference of change [95% confidence interval (CI)]: female = 1.0, 95% CI (0.15 to 1.8) p = 0.0211; male = −0.27, 95% CI (−1.6 to 1.06) p = 0.6629 (Fig. 2).
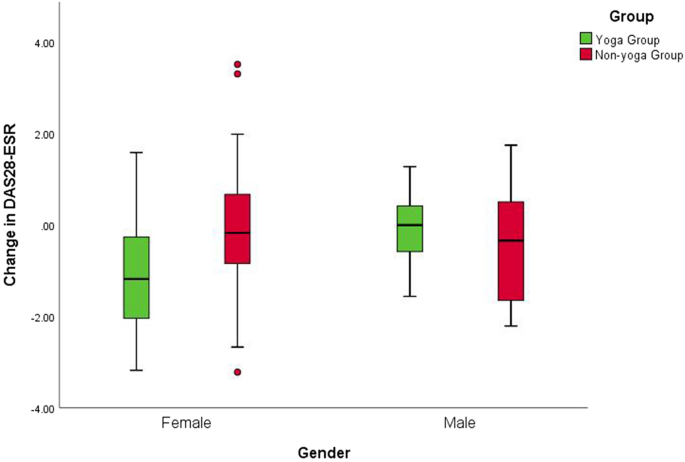
Gender interactions for change in disease activity after the intervention. Mean change of DAS28-ESR score with 95% CI for males in the yoga and the control groups; p = 0.6629 for between-group differences of change in the study, adjusted for baseline value. Mean change of DAS28-ESR score with 95% CI for females in the yoga and the control groups; p = 0.0211* for the between-group difference of change on the study, adjusted for baseline value.
Post-intervention differences in disease activity
There was a significant reduction in DAS28-ESR scores among the participants of yoga group [0.49, 95% (0.13 to 0.85)] with a significant interaction of group and time (ηp2 = 4.3; p = 0.042) (Table 2).
Post intervention differences in molecular markers
T cell subsets
The representative graphics from flow cytometry for Th17 and Treg cells measurements are depicted in Supplementary Fig. 1 and 2 respectively. Flow cytometry was performed to evaluate the Th17/Treg cell dynamics and aged T cell populations in both the groups. The mean percentage of Th17 cells (CD3+CD4+IL17+RORγt+ T cells) has shown a significant overall decline in yoga group (p = 0.004) from baseline to 8th week [1.7 ± 0.7 vs 0.9 ± 0.4; p < 0.0001], whereas a non-significant difference in non-yoga group from baseline to 8th week [1.5 ± 0.8 vs 1.7 ± 1.1; p = 0.381] (Fig. 3). Conversely, the mean percentage of Treg cells (CD3+CD4+CD25+CD127–Foxp3+ T cells) has shown a significant overall elevation in yoga group (p < 0.0001) from baseline to 8th week [1.1 ± 0.7 vs 2.1 ± 0.6; p < 0.0001], whereas a non-significant decline in non-yoga group from baseline to 8th week [1.4 ± 0.4 vs 1.1 ± 0.7; p = 0.093] (Fig. 3).
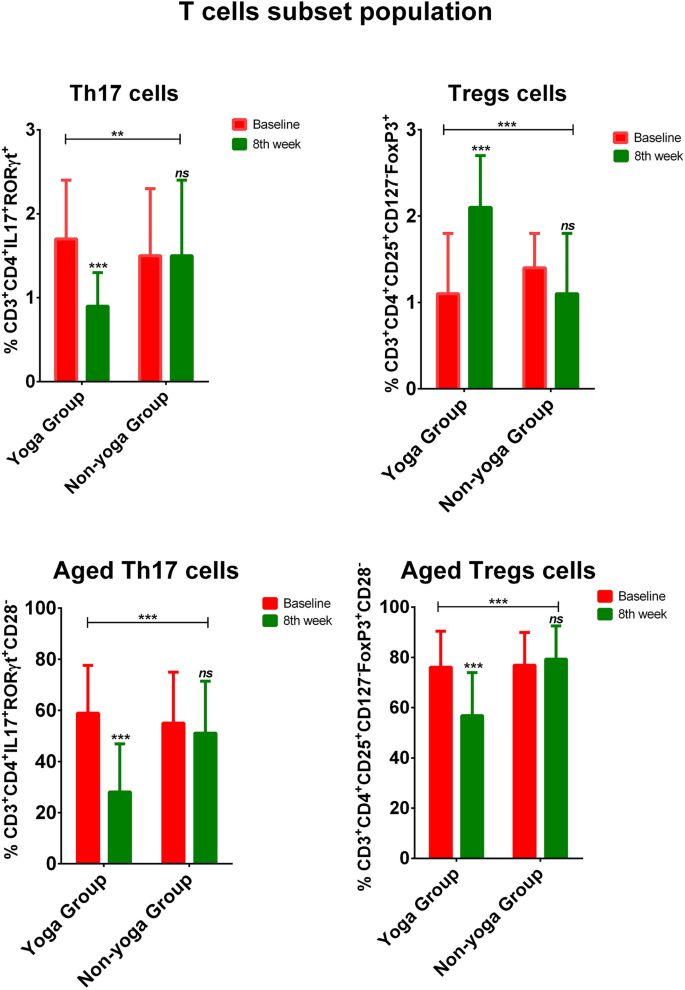
Frequency of Th17, Treg, aged Th17, and aged Treg cells in the yoga group and non-yoga group [p value (ns = p > 0.05; *p ≤ 0.05; **p ≤ 0.01;***p ≤ 0.001)].
Markers of immune aging
A significant overall decline in the mean percentage of aged Th17 cells (CD3+CD4+IL17+RORγt+CD28− T cells) was observed in the yoga group (p < 0.0001), from baseline to 8th week [58.9 ± 18.7 vs. 28.1 ± 18.8; p < 0.0001], whereas a non-significant difference in the non-yoga group from baseline to 8th week [55.0 ± 19.9 vs. 51.1 ± 20.3; p = 0.406] (Fig. 3). Similarly, a significant overall decline in the mean percentage of aged Treg cells (CD3+CD4+CD25+CD127−Foxp3+CD28− T cells) was seen in the yoga group (p < 0.0001) from baseline to 8th week [76.1 ± 14.3 vs. 56.8 ± 17.1; p < 0.0001], with no significant alterations observed in the non-yoga group from baseline to 8th week [76.9 ± 13.0 vs. 79.3 ± 13.2; p = 0.329] (Fig. 3).
Inflammatory markers
There were significant changes observed in various inflammatory markers after 8-weeks of intervention in yoga group as compared to non-yoga group. Pro-inflammatory cytokines like IL-6 [0.54, 95% CI (0.22 to 0.85); ηp2 = 10.4; p = 0.002], IL-17 [−0.96, 95% CI (−7.0 to 5.06); ηp2 = 33.4; p < 0.001] showed a significant decline with respect to interaction of group and time; whereas anti-inflammatory cytokines like TGF-β [−0.4, 95% CI (−0.7 to −0.07); ηp2 = 12.3; p = 0.001] and IL-10 [−13.5, 95% CI (−18.3 to −8.8); ηp2 = 45.7; p < 0.001] showed a significant increase in yoga group as compared to control group with respect to interaction of group and time (Table 2). Also, the change in mean within non-yoga group showed significantly increased levels of IL-17 [−18.4 ± 21.2; p < 0.001] compared to baseline levels (Table 2).
Epigenetic alterations
The percentage of global 5-mC was significantly higher in the yoga group as compared to the non-yoga group [−0.72, 95% CI (−1.3 to −0.16); ηp2 = 12.8; p = 0.001] (Table 2). Conversely, the percentage of global 5-hmC was significantly reduction in the yoga group after 8-weeks of intervention with a significant mean difference observed by group analysis [0.008, 95% CI (0.003 to 0.014); ηp2 = 4.4; p = 0.04)] (Table 2). Also, the HDAC1 levels were found to be significantly reduced after 8-weeks of intervention in the yoga group as compared to the non-yoga group [0.668, 95% CI (−0.32 to 1.6); ηp2 = 13.9; p < 0.001)] (Table 2).
Gene expression levels
The results for expression analysis showed significant downregulation in relative mRNA expression levels of RORγt (p = 0.0002), IL-17 (p = 0.0219), IL-6 (p = 0.0003), CXCL2 (p = 0.0065), and CXCR2 (p = 0.0284) in the yoga group as compared to the non-yoga group. The mean axis fold change of these transcripts was as follows in the yoga vs. non-yoga group: RORγt (−1.64 vs. 1.74), IL-17 (−3.42 vs. 3.15), IL-6 (−2.34 vs. 0.72), CXCL2 (−2.50 vs. 0.19), and CXCR2 (−4.90 vs. 1.32) respectively. The mRNA expression levels of JUN were not found to be different statistically (p = 0.7237) with a mean axis fold change of 1.05 vs. 1.38 in the yoga vs. non-yoga group. In the yoga group, the mRNA expression levels were significantly upregulated with the mean axis fold change of FoxP3 (1.93 vs. −0.65; p = 0.0169) and TGF-β (3.32 vs. 0.72; p = 0.0001) in the yoga vs. non-yoga group respectively (Fig. 4).

The relative mRNA expression levels of dysregulated transcripts in the yoga group and non-yoga group [p value (ns = p > 0.05; *p ≤ 0.05; **p ≤ 0.01;***p ≤ 0.001)].
Discussion
Our study is the first to highlight the positive impact of yoga on modulation of the T cell subsets, T cell aging markers, epigenetic alterations and associated transcription factors in RA. We found that 8 weeks of yoga practice significantly reduced disease activity, normalized the biomarkers associated with inflammation, and maintained Th17/Treg cell homeostasis. Further, yoga reduced the rate of immunological aging as seen by the reduction in the aged Th17 cell population (CD3+ CD4+ IL17+ RORγt+ CD28-T cells) and aged Treg cell population (CD3+ CD4+ CD25+ CD127-Foxp3+ CD28-T cells). Our findings suggest that yoga positively modified epigenetic changes such as global methylation levels, global hydroxyl methylation levels, and HDAC1 levels which may regulate gene expression patterns. The yoga group showed the downregulation of RORγt, IL-17, IL-6, CXCL2, CXCR2, and upregulation of FoxP3 and TGF-β. These findings suggest that yoga possesses an immune-modulatory potential which induces molecular remission in RA by influencing its pathobiology at both cellular and molecular level.
In the present study, the impact of regular practice of yoga was beneficial as there was a significant reduction in the systemic levels of pro-inflammatory markers (IL-6 and IL-17) and various transcripts associated with pro-inflammatory cytokines. IL-6 is essential for the development of RA’s systemic(lung, heart skin, brain) and joint inflammation, immune system abnormalities, and joint swelling30. The systemic inflammatory signs and symptoms of RA, which are mediated by IL-6, include fever, malaise, sleep disturbances, muscular weakness, and anemia31. In order to promote the production of adhesion molecules and draw leukocytes to the affected joints, IL-6 induces endothelial cells to secrete IL-8 and monocyte chemoattractant protein-1 (MCP-1) locally at joint level32. IL-6 can promote osteoclast differentiation and synoviocyte proliferation by activation of the NF-kappa B receptor activator ligand (RANKL). Through STAT3-mediated activation of the RORγt in the presence of TGF-β, IL-6 stimulates the differentiation of Th17 cells while inhibiting TGF-induced Treg cell differentiation33. Thus, IL-6 encourages Th17 over Treg dominance in the effector CD4+ T cell subsets, which is regarded to be a key factor in the emergence of RA and other immune-mediated illnesses30. Another important inflammatory marker, IL-17 affects RA joints by promoting the development of matrix metalloproteases and pro-inflammatory cytokines, which contribute to tissue inflammation and destruction34. Th17 cells play a unique role in immune activity through the generation of effector cytokines such as IL-17A, IL-17F, IL-22, etc. Th17 cells influence the development of osteoclasts and bone resorption by the release of IL-17, an activate monocytes to create pro-inflammatory cytokines, accelerating the inflammatory cascade35,36. Expression of CXCL2 was significantly increased in RA patients and was associated with bony erosions37. CXCL2 is a pro-inflammatory factor which is associated with biological signs of progress, such as angiogenesis, inflammation and cancer. CXCL2 stimulated osteoclastogenesis via extracellular receptor kinase (ERK) mitogen‐activated protein kinase (MAPK) and nuclear factor kappa B pathways37. Leukocyte trafficking is regulated by chemokines both during homeostasis and immunological responses. CXCR2 plays a cell-autonomous function in the migration of neutrophils to inflamed joints, which is essential for the emergence of auto-immune arthritis38. Furthermore. our study demonstrates that yoga practice down-regulates the mRNA transcript levels of pro-inflammatory genes (RORγt, IL-6, IL-17, CXCL2, CXCR2), hence reducing systemic and local inflammation in RA, thus overall providing a clear clinical and molecular evidence of the impact of yoga on RA. Yoga, is mind body energy medicine, comprises of both psychological and physical components and is thus ideal as an adjunct management system of this severe painful disabling autoimmune arthritis like RA and has been documented from previous studies from our group for its role as a powerful adjunct to modern medicine in disease management, rehabilitation and promotion of health.
Yoga is a centuries-old method of unifying the mind, body, and soul. The Patanjali’s ashtanga (eight limb) Yoga includes yama (abstinences), niyama (observances), asana (yoga postures), pranayama (breath control), pratyahara (withdrawal of the senses), dharana (concentration), dhyana (meditation) and samadhi (absorption)18. Dharna, dhyana and samadhi are antaranga yoga. Yoga originated from the belief that stress is the cause of all illnesses, which gives the practice advantages for both physical and mental health39. Stress impacts every cell of the body and every organ system and yoga thus targets both the mind and body reducing stress by building emotional resilience by promoting neuroplasticity. Not only is there increase in several neurochemicals like brain derived neurotrophic factor (BDNF), dehydroepiandrosterone (DHEA), melatonin, serotonin but also actual anatomical changes in the brain in the hippocampus, prefrontal cortex which show increase in gray matter and concomitant decrease in size of amygdala. IL-10 possesses anti-inflammatory and immunoregulatory properties. Yoga has the potential to mediate the inflammatory pathways by increasing the levels of anti-inflammatory mediators and reducing the pro-inflammatory markers19,40. IL-10 prevents T-cell responses to specified antigens and inhibits the synthesis of proinflammatory cytokines and chemokines41. The main mechanism by which IL-10 exerts its effects is by inhibiting the costimulatory capabilities of macrophages and promoting the proliferation and differentiation of antibody-forming B cells. The effectiveness of IL-10 in reducing inflammation and autoreactivity has been demonstrated in preclinical research using a range of animal models, including collagen-induced arthritis42. TGF-β is a regulatory cytokine which promotes the growth of Treg cells and controls CD4+ T cell polarization. Depending on the cytokine environment, TGF- β stimulates the differentiation of Th17 and Treg cells43. The transcription factor RORγt, which is the principal regulator of Th17 cells, is activated and controlled in its expression by TGF- signaling and STAT344. TGF-β can induce the differentiation of CD4+ T cells into Th17 cells in the presence of IL-6 or IL-21. Our results have shown that the yoga group showed significant upregulation of mRNA expression levels of FoxP3 and TGF-β as compared to the non-yoga group, further confirming the immune-modulatory role of yoga. The dimers of members of the Jun, Fos and activating transcription factor protein families make up the transcription factor activator protein 1 (AP-1)45. Jun mostly acts as a proliferating growth regulator of cells46.In the present study, the mRNA expression levels of JUN had shown a declining trend in the yoga group; however, the difference was not statistically significant as compared to the non-yoga group.
The immune dysregulation in RA is attributed to increased secretion of inflammatory cytokines by effector Th17 cells and a loss of Treg cells suppressor function. Yoga’s anti-inflammatory properties work to restore immune homeostasis to its ideal state and promote natural immunological tolerance to treat autoimmune diseases20,21. Our results also showed that regular practice of yoga was associated with increased Treg cell population and decreased Th17 cell population. Our findings imply that yoga-induced variations in serum cytokine levels stimulate changes in Treg and Th17 cell populations, which may shift the delicate immune system balance to reestablish tolerance. A study on the effect of a 12-week program of regular tai chi chuan exercise showed beneficial effects on functional mobility, beliefs about the benefits of exercise on physical and psychological health and maintained immune regulation in middle-aged volunteers by increase in their Treg cells47,48. Our results were in concordance with a study conducted on the ischemic cardiomyopathy rat model in which the Th17/Treg ratio was much lower in the rats trained on treadmill (Slope 0°, 12 m/min, 30 min/time) five times per week, for 12 weeks than non-training group rats49.Few more studies focused on the impact of endurance training and physical exercise on athletes where a diversion in the Th17/Treg cell balance was seen49,50. Our study has documented a post-yoga reduction in the mean percentage of Th17 cells (CD3+ CD4+ IL17+ RORγt+ T cells) in yoga group from baseline to 8th week whereas the mean percentage of Treg cells (CD3+ CD4+ CD25+ CD127− Foxp3+ T cells) has shown a significant elevation in yoga group. RA patients experience the signs of accelerated ageing like reduced thymic functioning, expansion of late-differentiated effector T cells, increased telomeric attrition, and excessive production of cytokines (senescence-associated secretory phenotype)12. Early onset of age-related co-morbidities like osteoporosis, cardiovascular issues, and cognitive decline have all been linked to the progression of RA51. The crucial co-stimulatory protein CD28 is expressed on undifferentiated CD4+ and CD8+ T lymphocytes. Since the MHC-I-bound peptide antigen has a low affinity, activating T cells using antigen-presenting cells (APCs) alone is not enough to fully activate and sustain T cells52. To extend T cell responses, the co-stimulatory signal from the CD28 cell surface receptor that binds with CD86 or CD80 on antigen presenting cells is required. Yoga possesses the potential to reduce the pace of immunological aging which can be attributed to the reduced mean percentage of aged Th17 cells (CD3+ CD4+ IL17+ RORγt+ CD28-T cells) as well as aged Treg cells (CD3+ CD4+ CD25+ CD127− Foxp3+ CD28-T cells) after 8 weeks of practice.
Inflammatory changes may be influenced by epigenetic mechanisms that subsequently lead to gene expression alterations and, eventually, protein expression53. The epigenome controls gene expression and is susceptible to changes brought on by stress, lifestyle changes and other environmental variables54. The most extensively researched epigenetic modification is DNA methylation, which is increasingly recognized as a meaningful biomarker in the pathogenesis of RA55. High levels of inflammation and poor physical health have both been linked to changes in DNA methylation56. Since epigenetic modifications have the capacity to be reversed, they can be used to gauge how well clinical treatments are working. Recent studies have reported a global DNA hypomethylation in immune cells of RA patients as compared to healthy individuals57. Global DNA hypomethylation is implicated in various inflammatory and autoimmune disorders and leads to aberrant gene expression58. In RA, the low levels of methylation at specific CpG sites in fibroblast-like synoviocytes is correlated with overexpression of genes which are important for the disease pathogenesis such as growth factors/ receptors, extracellular matrix proteins, adhesion molecules, and matrix degrading enzymes, etc59,60. In our study, we have seen an overall increase in the percentage of global 5-mC DNA in yoga group over non-yoga group after 8-weeks of yoga intervention, which might lead to the downregulation of pro-inflammatory genes like RORγt, IL-6, IL-17, CXCL2, CXCR2, etc. DNA demethylation also occurs via oxidative modification followed by removal of the base61. Active DNA demethylation occurs due to the presence of 5-hmC. In order to study the effect of yoga on the status of DNA demethylation in RA patients, the global 5hmC percentage was estimated. This is the first study to document an overall decrease in the percentage of global 5-hmC DNA in yoga group over non-yoga group after 8- weeks of yoga intervention. Modifications of histones are major epigenetic markers that regulate gene expression and establish various cell phenotypes62. The open structure of chromatin makes the chromatin available to transcription factors and can increase gene expression dramatically. Histone acetylation is important for gene expression regulation, while histone deacetylation contributes to chromatin condensation and gene transcription repression63. Inhibition of HDAC function by epigenetic or non-epigenetic mechanisms can lead to the development of RA and other autoimmune inflammatory diseases, affecting the complex modulation of intracellular signaling pathways64. HDAC inhibitors exhibit anti-arthritic activities through modulation of only a small percentage of gene expression involved in chronic inflammation and apoptosis65. HDAC inhibitors act via epigenetic and non-epigenetic processes to generate immunomodulatory effects. HDAC inhibitors demonstrated anti-inflammatory effects in animal models of arthritis and synovial tissues in RA patients66. In our study, we have seen a reduction in the levels of HDAC1 in yoga group as compared to non-yoga group after 8-weeks of yoga. The effects of HDAC inhibitors on cellular activation have been attributed to epigenetic regulation, signal transduction modifications, and gene expression modulation via regulation of mRNA stability.
Lack of an active control group was one of the study’s shortcomings because the non-yoga group had no equal attention control intervention and only received medication therapy in comparison to the active yoga intervention group. Therefore, including such a group would further rule out the therapeutic results that were specifically linked to the yoga intervention. In both the yoga and non-yoga groups, there were fewer men, which was explained by the fact that women had a higher prevalence of RA than men (3:1). It is challenging to maintain a regular schedule for a long-term RA management or home practice regimen because each session of yoga lasted for 120 min every day while being supervised by a certified yoga instructor. Also, the lack of long-term follow-up of the participants made it difficult to predict how quickly participants returned to their baseline levels. In order to investigate the long-term advantages of the yoga practice, we intend to conduct additional research with a large sample size and long-term follow-ups and practice of yoga for shorter duration.
In conclusion, our study findings highlight that yoga possesses an immune-modulatory potential which induces molecular remission and reestablishes immunological tolerance in RA by influencing its pathobiology by optimizing inflammatory markers, maintaining immune-homeostasis, reducing the rate of immune-aging and improving RA health outcome. The 8 weeks of yoga practice significantly reduces disease activity, maintains Th17/Treg cell homeostasis and reduces inflammatory processes by optimizing the levels of various pro-inflammatory cytokines, and anti-inflammatory cytokines with changes in gene expression patterns. Yoga positively modifies the epigenome by elevating global methylation levels, reducing global hydroxyl methylation levels, and HDAC levels which may cause the normalization of dysregulated gene expression. Hence, yoga can be used as an adjuvant therapy in RA as it boosts physical functioning, enhances psychological wellbeing and reestablishes immunological tolerance.
Methods
Ethics declarations and study design
This study was a prospective, single-blinded, randomized controlled trial with active RA patients, aimed at analyzing the effects of an 8-week yoga practice in RA patients on standard drug therapy. The study was initiated after obtaining ethical clearance (IECPG-211/24.02.2016) from the Institute Ethics Committee of AIIMS, New Delhi, India, and registration under the clinical trials registry, India (CTRI/2017/05/008589, Registered on 17.05.2017). All methods were performed in accordance with the relevant guidelines and regulations. All the participants gave written informed consent before the study protocol’s commencement.
Participants and eligibility criteria
The study participants were recruited from the outpatient unit of Rheumatology department of AIIMS, New Delhi. RA patients, 18–60 years old, diagnosed as per 2010 ACR/EULAR RA classification criteria26, whose DAS28ESR was > 2.6 and who were on standard medical treatment for at least 6 months were recruited after obtaining written informed consent. Patients with any other overlapping autoimmune diseases, chronic systemic diseases, history of administration of oral or intra-articular steroids in the previous six months, medical treatment for any other illnesses, intake of any form of herbo-mineral, antioxidant, homeopathic, or ayurvedic supplementation (Boswellia serrata, Ricinus communis, dry ginger powder, fenugreek seeds, curcumin, ashwagandha etc.) or already practicing yoga and meditation in any way were excluded from the study. Also, during the execution of the study, the events which resulted in the exclusion were the inability to comply with regular yoga and deviating from their usual lifestyle habit.
Sample size calculation
The sample size calculation for the study assumed to detect a standardized effect size [difference in mean change in DAS28-ESR between the two groups/pooled standard deviation (SD)] of 0.8 with a 95% confidence level and 80% power, considering the mean and SD of a previous study by Evans et al. (2011)27. Considering some loss to follow-up, we enrolled a total of 64 patients in the study and randomized into two groups—yoga group (32 patients) and non-yoga group (32 patients).
Randomization
For this investigator blinded study, sequentially labeled sealed opaque envelopes were used to conceal random numbers as described earlier20. As each patient was enrolled, they were assigned to the next numbered envelope and the associated group, either yoga group or non-yoga group. The sequence of random numbers was generated by permuted block randomization of variable block size with the assistance of the web tool research randomizer (https://www.randomizer.org/). The participants were not blinded to the study, whereas investigators who interviewed patients, conducted experiments, and performed statistical analyses blinded to the group status of the patients.
Intervention
The participants were randomized into two groups- the yoga group and non-yoga group. All the study participants were asked to undergo a clinical evaluation and provide a blood sample on day 0 (baseline) and the end of 8th week of intervention.
Yoga program (yoga group)
As described previously19, the participants of the yoga group were administered a standardized yoga program for eight weeks, which was suitable for active RA patients, so that it did not cause any further irritation to already inflamed joints. The patients with deformed joints and limitations were advised to undergo customized physical postures, relaxation exercises, and practiced breathing forming a slow and deep breathing pattern with exhalation being longer than inhalation. The comprehensive yoga program incorporated the components of Patanjali’s ashtanga yoga. Briefly, the intervention comprised of yogic practices including asanas (physical postures), pranayama (regulated breathing techniques), dhyana (meditation), and savasana (relaxation techniques), followed by interactive counseling sessions on yoga, stress management, nutrition, as well as personal lifestyle management (Supplementary Fig. 3). Yoga program was administered five times a week for 120 min duration per session for 8 weeks19,25. The sessions were conducted by the registered and well-qualified yoga instructor at the Laboratory for Molecular Reproduction & Genetics, Department of Anatomy, AIIMS, New Delhi. Adherence was monitored with participant diary and yoga teacher’s remarks at each visit. As such, there was no home regimen to be followed for the yoga practice due to the extensive time commitment of each session. The patients were encouraged to incorporate yoga into their lifestyle after the end of the 8 weeks of intervention. The patients of this group continued with their standard disease-modifying anti-rheumatic drugs (DMARDs) as per the prescription of rheumatologists.
Usual care control (non-yoga group)
As described previously19, the patients assigned to the non-yoga group continued with their ongoing medical care, which included DMARDs prescribed by the rheumatologists. The patients followed their normal daily physical activities with no change in their daily lifestyle for eight weeks.
Outcome measures
Primary outcome
The disease activity of the patients was assessed by DAS28-ESR28. The DAS28-ESR consisted of four components: tender joint count, swollen joint count, visual analog scale (VAS) score of the patient’s global health, and ESR. In DAS28-ESR, a rating of ≤ 2.6 represents remission, > 2.6 to 3.2 represents low disease activity, > 3.2 to 5.1 represents moderate disease activity, and ≥ 5.1 represents high disease activity.
Change in disease severity was measured by disease activity score—erythrocyte sedimentation rate (DAS28-ESR) from baseline (day 0) to 8-weeks and recorded as primary outcome.
Secondary outcome
All parameters were evaluated on day 0 (baseline) and 8th week (follow-up) of the intervention and the fasting blood samples were obtained at 8 am in the morning.
Alterations in cellular and molecular markers including: (a) T cell subset population: Th17 (CD3+ CD4+ IL17+ RORγt+) cells and Treg (CD3+ CD4+ CD25+ CD127− Foxp3+) cells, (b) markers of immune aging: aged Th17 (CD3+ CD4+ IL17+ RORγt+ CD28−) cells and aged Treg (CD3+ CD4+ CD25+ CD127− Foxp3+ CD28−) cells, (c) inflammatory markers: IL-6, IL-17, TGF-β, and IL-10 (d) epigenetic alterations: 5-methyl cytosine, 5-hydroxymethyl cytosine and HDAC1, and (e) gene expression levels: RORγt, FoxP3, IL-17, IL-6, TGF-β,CXCL2,CXCR2, and JUN were documented.
Measurement of molecular markers
Following techniques were employed for the measurement of molecular markers:
Phenotyping of T cell subsets by flow cytometry
The isolated PBMCs (1 × 106 cells /ml) were stained with a panel of fluorochrome-conjugated monoclonal antibodies (Thermo Fisher Scientific, USA) for the surface markers, namely CD3, CD4, CD28, CD25, and CD127 and intra-cellular cytokines/transcription factors, namely IL-17, RORγt, and FoxP3. Alexa Fluor 488-labeled anti-human CD3, PerCP Cy5.5-labeled anti-human CD4, APC- labeled anti-human IL-17, PE-labeled anti-human RORγt were used to examine CD3+CD4+IL17+RORγt+ T cells. Alexa Fluor 488-labeled anti-human CD3, PerCP Cy5.5-labeled anti-human CD4, PE-labeled anti-human CD25, APC-eFluor 780-labeled anti-human CD127, and APC-labeled anti-human FoxP3 were used to detect CD3+CD4+CD25+CD127−Foxp3+ T cells. PE-Cyanine7-labeled anti-human CD28 was used for aged T cell population i.e. aged Th17 cells(CD3+CD4+IL17+RORγt+CD28− T cells) and aged Treg cells (CD3+CD4+CD25+CD127–Foxp3+CD28− T cells).
The isolated PBMCs (1 × 106 cells /ml) were taken in FACS tubes for immuno-staining. The recommended value of monoclonal antibodies (for surface markers as mentioned above) were added, mixed thoroughly, and incubated on ice or 4 °C for 45 min in the dark. Cells were washed with staining buffer (1× PBS with 1% FBS) and pelleted at 1200 rpm for 10 min. For intracellular staining, cells were permeabilized and added with permeabilization buffer (Thermo Fisher Scientific, USA). The cell pellet was resuspended in 100 μl of transcription factor staining buffer (Thermo Fisher Scientific, USA), and with the recommended value of monoclonal antibodies (for transcription factors & intracellular cytokines) and incubated in the dark at room temperature for 1 h. After two steps of washing with PBS, the cells were resuspended in 100 μl of with permeabilization buffer (Thermo Fisher Scientific, USA) and fixed by adding 20 μl of 4% paraformaldehyde (PFA) and stored at 4 °C. Fixed cells were flow- cytometrically acquired & analyzed by BD Fortessa X20 flow cytometer (BD Biosciences, San Jose, CA) equipped with FACSDIVA™ software. Fifty thousand events were acquired per sample within a typical forward, and side scatter gate set to exclude dead cells and debris. Data were analyzed using FlowJo 7.22 (Tree Star Ashland, Orlando, FLA) software.
Detection of inflammatory and epigenetic markers
Serum levels of IL-6 (Gen-Probe, Diaclone Diagnostic, France), IL-17 (Gen-Asia Biotech, China), IL-10 (Bioassay Tech Laboratory, China), and HDAC1 (Qayee Bio-Technology) were estimated by ELISA using commercially available kits. Briefly, the workflow of the HDAC1 ELISA kit utilizes the sandwich ELISA methodology where the plate was pre-coated with human HDAC1 antibody. The serum samples were then added onto the pre-coated wells. And, then biotinylated human HDAC1 antibody was added, which binded to HDAC1 in the sample. After the addition of Streptavidin-HRP developer followed by substrate and stop solution, the well absorbance for HDAC1 was measured at 450 nm. The 5-methylcytosine DNA ELISA kit (Enzo Life Sciences, Inc., USA) and 5-hydroxymethyl cytosine DNA ELISA kit (Enzo Life Sciences, Inc., USA) was used to quantify the percent 5-mC DNA and 5-hmC DNA respectively. Briefly, the workflow for the 5-mC DNA ELISA kits utilize the indirect ELISA methodology where 100 ng denatured, single-stranded DNA (ssDNA) samples per well were coated on the plate wells and a 5-mC mAb and conjugate HRP-Ab were then added to the wells. The detection of 5-mC was done after the addition of the HRP developer by measuring well absorbance at 405–450 nm. Briefly, the workflow of the 5-hmC DNA ELISA kit utilizes the sandwich ELISA methodology where a 5-hmC pAb was coated to the bottom of plate well surfaces. The ssDNA (100 ng/well) sample was then added onto the well surface which binded to 5-hmC pAb’s and was then recognized by a conjugate DNA HRP-Ab. After the addition of HRP developer, the well absorbance for 5-hmC DNA was measured at 405–450 nm. Serum TGF-β levels were estimated by a magnetic bead-based multiplex assay using Bio-Plex Pro TGF-β Assays (Bio-Rad Laboratories Inc., USA) according to the manufacturer’s guidelines. Quality-control assays for biomarkers and validation were performed.
Detection of gene expression patterns
As described earlier20, total RNA was isolated from 1 ml of freshly obtained EDTA blood by TRIzol manual method followed by complementary DNA (cDNA) synthesis by reverse transcribing 1000 ng of RNA by using the iScript cDNA synthesis kit (BioRad). The CFX96 realtime system (BioRad, CA, United States) quantified the relative gene expression using Brilliant III UltraFast SYBR Green qPCR Master Mix. The protocol for gene amplification was standardized at 35 cycles. Normalization of the amount of expressed mRNA used two internal housekeeping genes 36B4 and β-actin. The relative fold of gene expression was done by the 2−ΔΔCt method29. The primer sequences for the genes used in the study are shown in Table 3.
Statistical analysis
All statistical analyses were carried out on an intent-to-treat basis with the baseline observation carried forward approach using IBM SPSS Statistics for Macintosh, Version 25.0. (IBM Corp. Armonk, NY, United States) and GraphPad Prism Version 6.01. (GraphPad Software, Inc., San Diego, CA). Chi-square test and Fisher’s exact test compared the baseline characteristics between the two groups. The assessment of interaction effects among baseline parameters was carried out by mixed factorial design ANOVA. For within-group analysis, paired t-test was used to study the difference between pre- to post-intervention for normally distributed data, or Wilcoxon signed-rank tests for continuous variables without normal distribution. For between-group analysis, the repeated measure ANOVA was used to study the intervention effects along with the interaction of time and group. A p-value of < 0.05 was considered to be statistically significant.
Data availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
References
-
Tobón, G. J., Youinou, P. & Saraux, A. The environment, geo-epidemiology, and autoimmune disease: Rheumatoid arthritis. J. Autoimmun. 35, 10–14 (2010).
-
Dejaco, C., Duftner, C., Grubeck-Loebenstein, B. & Schirmer, M. Imbalance of regulatory T cells in human autoimmune diseases. Immunology 117, 289–300 (2006).
-
De Almeida, D. E. et al. Immune dysregulation by the rheumatoid arthritis shared epitope. J. Immunol. Baltim. Md 1950(185), 1927–1934 (2010).
-
Yap, H.-Y. et al. Pathogenic role of immune cells in rheumatoid arthritis: Implications in clinical treatment and biomarker development. Cells 7, 161 (2018).
-
Afzali, B., Lombardi, G., Lechler, R. I. & Lord, G. M. The role of T helper 17 (Th17) and regulatory T cells (Treg) in human organ transplantation and autoimmune disease. Clin. Exp. Immunol. 148, 32–46 (2007).
-
Martinez, G. J., Nurieva, R. I., Yang, X. O. & Dong, C. Regulation and function of proinflammatory TH17 cells. Ann. N. Y. Acad. Sci. 1143, 188–211 (2008).
-
Corthay, A. How do regulatory T cells work?. Scand. J. Immunol. 70, 326–336 (2009).
-
Capone, A. & Volpe, E. Transcriptional regulators of T helper 17 cell differentiation in health and autoimmune diseases. Front. Immunol. 11, 348 (2020).
-
Pereira, L. M. S., Gomes, S. T. M., Ishak, R. & Vallinoto, A. C. R. Regulatory T cell and forkhead box protein 3 as modulators of immune homeostasis. Front. Immunol. 8, 605 (2017).
-
Jiang, Q., Yang, G., Liu, Q., Wang, S. & Cui, D. Function and role of regulatory T cells in rheumatoid arthritis. Front. Immunol. 12, 626193 (2021).
-
Lee, G. R. The balance of Th17 versus Treg cells in autoimmunity. Int. J. Mol. Sci. 19, 730 (2018).
-
Weyand, C. M., Yang, Z. & Goronzy, J. J. T cell aging in rheumatoid arthritis. Curr. Opin. Rheumatol. 26, 93–100 (2014).
-
Chalan, P., van den Berg, A., Kroesen, B.-J., Brouwer, L. & Boots, A. Rheumatoid arthritis, immunosenescence and the hallmarks of aging. Curr. Aging Sci. 8, 131–146 (2015).
-
Goronzy, J. J. & Weyand, C. M. Mechanisms underlying T cell ageing. Nat. Rev. Immunol. 19, 573–583 (2019).
-
Shao, L. et al. Deficiency of the DNA repair enzyme ATM in rheumatoid arthritis. J. Exp. Med. 206, 1435–1449 (2009).
-
Li, Y. et al. Deficient activity of the nuclease MRE11A induces T cell aging and promotes arthritogenic effector functions in patients with rheumatoid arthritis. Immunity 45, 903–916 (2016).
-
Belk, J. A., Daniel, B. & Satpathy, A. T. Epigenetic regulation of T cell exhaustion. Nat. Immunol. 23, 848–860 (2022).
-
Gautam, S. & Dada, R. Molecular mechanisms underlying the effects of yoga. In Handbook of Research on Evidence-Based Perspectives on the Psychophysiology of Yoga and Its Applications. 103–123. https://www.igi-global.com/chapter/molecular-mechanisms-underlying-the-effects-of-yoga/www.igi-global.com/chapter/molecular-mechanisms-underlying-the-effects-of-yoga/261146. https://doi.org/10.4018/978-1-7998-3254-6.ch007 (2021).
-
Gautam, S., Tolahunase, M., Kumar, U. & Dada, R. Impact of yoga-based mind-body intervention on systemic inflammatory markers and co-morbid depression in active rheumatoid arthritis patients: A randomized controlled trial. Restor. Neurol. Neurosci. 37, 41–59 (2019).
-
Gautam, S., Kumar, M., Kumar, U. & Dada, R. Effect of an 8-week yoga-based lifestyle intervention on psycho-neuro-immune axis, disease activity, and perceived quality of life in rheumatoid arthritis patients: A randomized controlled trial. Front. Psychol. 11, 2259 (2020).
-
Gautam, S., Kumar, U. & Dada, R. Yoga and its impact on chronic inflammatory autoimmune arthritis. Front. Biosci. Elite Ed. 13, 77–116 (2021).
-
Arora, S. & Bhattacharjee, J. Modulation of immune responses in stress by yoga. Int. J. Yoga 1, 45–55 (2008).
-
Falkenberg, R. I., Eising, C. & Peters, M. L. Yoga and immune system functioning: A systematic review of randomized controlled trials. J. Behav. Med. 41, 467–482 (2018).
-
Gautam, S., Kumar, U., Kumar, M., Rana, D. & Dada, R. Yoga improves mitochondrial health and reduces severity of autoimmune inflammatory arthritis: A randomized controlled trial. Mitochondrion 58, 147–159 (2021).
-
Gautam, S., Kumar, U., Mishra, R. & Dada, R. HLA-G 3’UTR polymorphisms & response to a yoga-based lifestyle intervention in rheumatoid arthritis: A randomized controlled trial. Indian J. Med. Res. 155, 253–263 (2022).
-
Kay, J. & Upchurch, K. S. ACR/EULAR 2010 rheumatoid arthritis classification criteria. Rheumatol. Oxf. Engl. 51(Suppl 6), vi5-9 (2012).
-
Evans, S. et al. A randomized controlled trial examining Iyengar yoga for young adults with rheumatoid arthritis: a study protocol. Trials 12, 19 (2011).
-
Nishimoto, N. & Takagi, N. Assessment of the validity of the 28-joint disease activity score using erythrocyte sedimentation rate (DAS28-ESR) as a disease activity index of rheumatoid arthritis in the efficacy evaluation of 24-week treatment with tocilizumab: subanalysis of the SATORI study. Mod. Rheumatol. 20, 539–547 (2010).
-
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods San Diego Calif. 25, 402–408 (2001).
-
Yoshida, Y. & Tanaka, T. Interleukin 6 and rheumatoid arthritis. BioMed Res. Int. 2014, 698313 (2014).
-
Cronstein, B. N. Interleukin-6—A key mediator of systemic and local symptoms in rheumatoid arthritis. Bull. NYU Hosp. Jt. Dis. 65(Suppl 1), S11-15 (2007).
-
Guo, Q. et al. Rheumatoid arthritis: Pathological mechanisms and modern pharmacologic therapies. Bone Res. 6, 15 (2018).
-
O’Brien, W. et al. Receptor activator of nuclear factor kappa-B (RANK) independent osteoclast formation and bone erosion in inflammatory arthritis. Arthritis Rheumatol. Hoboken NJ 68, 2889–2900 (2016).
-
Robert, M. & Miossec, P. IL-17 in rheumatoid arthritis and precision medicine: From synovitis expression to circulating bioactive levels. Front. Med. 5, 364 (2019).
-
Tesmer, L. A., Lundy, S. K., Sarkar, S. & Fox, D. A. Th17 cells in human disease. Immunol. Rev. 223, 87–113 (2008).
-
Shabgah, A. G., Fattahi, E. & Shahneh, F. Z. Interleukin-17 in human inflammatory diseases. Postepy Dermatol. Alergol. 31, 256–261 (2014).
-
Wang, X. et al. Increased expression of CXCL2 in ACPA-positive rheumatoid arthritis and its role in osteoclastogenesis. Clin. Exp. Immunol. 203, 194–208 (2021).
-
Jacobs, J. P. et al. Deficiency of CXCR2, but not other chemokine receptors, attenuates a murine model of autoantibody-mediated arthritis. Arthritis Rheum. 62, 1921–1932 (2010).
-
Woodyard, C. Exploring the therapeutic effects of yoga and its ability to increase quality of life. Int. J. Yoga 4, 49–54 (2011).
-
Vijayaraghava, A., Doreswamy, V., Narasipur, O. S., Kunnavil, R. & Srinivasamurthy, N. Effect of yoga practice on levels of inflammatory markers after moderate and strenuous exercise. J. Clin. Diagn. Res. JCDR 9, CC08-12 (2015).
-
Taylor, A., Verhagen, J., Blaser, K., Akdis, M. & Akdis, C. A. Mechanisms of immune suppression by interleukin-10 and transforming growth factor-β: The role of T regulatory cells. Immunology 117, 433–442 (2006).
-
Moore, K. W., de Waal Malefyt, R., Coffman, R. L. & O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 19, 683–765 (2001).
-
Zhang, S. The role of transforming growth factor β in T helper 17 differentiation. Immunology 155, 24–35 (2018).
-
Aarts, J. et al. Local inhibition of TGF-β1 signaling improves Th17/Treg balance but not joint pathology during experimental arthritis. Sci. Rep. 12, 3182 (2022).
-
Garces de losFayos Alonso, I. et al. The role of activator protein-1 (AP-1) family members in CD30-positive lymphomas. Cancers 10, 93 (2018).
-
Johnston, I. M. et al. Regulation of a multigenic invasion programme by the transcription factor, AP-1: Re-expression of a down-regulated gene, TSC-36, inhibits invasion. Oncogene 19, 5348–5358 (2000).
-
Oh, B. et al. The effects of Tai Chi and Qigong on immune responses: A systematic review and meta-analysis. Medicines 7, 39 (2020).
-
Wang, M.-Y. & An, L.-G. Effects of 12-Week’s Tai Chi Chuan Practice on the Immune Function of Female College Students Who Lack Physical Exercise. https://doaj.org/article/51b2fc58e04542e2b3224c586645a1b2 (2011).
-
Chen, Z. et al. Effect of aerobic exercise on Treg and Th17 of rats with ischemic cardiomyopathy. J Cardiovasc. Transl. Res. 11, 230–235 (2018).
-
Perry, C. et al. Endurance exercise diverts the balance between Th17 cells and regulatory T cells. PLoS ONE 8, e74722 (2013).
-
López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M. & Kroemer, G. The hallmarks of aging. Cell 153, 1194–1217 (2013).
-
Beyersdorf, N., Kerkau, T. & Hünig, T. CD28 co-stimulation in T-cell homeostasis: A recent perspective. ImmunoTargets Ther. 4, 111–122 (2015).
-
Bayarsaihan, D. Epigenetic mechanisms in inflammation. J. Dent. Res. 90, 9–17 (2011).
-
Turner, B. M. Epigenetic responses to environmental change and their evolutionary implications. Philos. Trans. R. Soc. B Biol. Sci. 364, 3403–3418 (2009).
-
Nemtsova, M. V. et al. Epigenetic changes in the pathogenesis of rheumatoid arthritis. Front. Genet. 10, 570 (2019).
-
Ramos-Lopez, O., Milagro, F. I., Riezu-Boj, J. I. & Martinez, J. A. Epigenetic signatures underlying inflammation: An interplay of nutrition, physical activity, metabolic diseases, and environmental factors for personalized nutrition. Inflamm. Res. 70, 29–49 (2021).
-
de Andres, M. C. et al. Assessment of global DNA methylation in peripheral blood cell subpopulations of early rheumatoid arthritis before and after methotrexate. Arthritis Res. Ther. 17, 233 (2015).
-
Jeffries, M. A. & Sawalha, A. H. Autoimmune disease in the epigenetic era: How has epigenetics changed our understanding of disease and how can we expect the field to evolve?. Expert Rev. Clin. Immunol. 11, 45–58 (2015).
-
Karouzakis, E., Gay, R. E., Michel, B. A., Gay, S. & Neidhart, M. DNA hypomethylation in rheumatoid arthritis synovial fibroblasts. Arthritis Rheum. 60, 3613–3622 (2009).
-
Karouzakis, E. et al. Analysis of early changes in DNA methylation in synovial fibroblasts of RA patients before diagnosis. Sci. Rep. 8, 7370 (2018).
-
Kriaucionis, S. & Heintz, N. The nuclear DNA base, 5-hydroxymethylcytosine is present in brain and enriched in Purkinje neurons. Science 324, 929–930 (2009).
-
Handy, D. E., Castro, R. & Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 123, 2145–2156 (2011).
-
Seto, E. & Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb. Perspect. Biol. 6, 18713 (2014).
-
Vojinovic, J. & Damjanov, N. HDAC inhibition in rheumatoid arthritis and juvenile idiopathic arthritis. Mol. Med. 17, 397–403 (2011).
-
Grabiec, A. M., Korchynskyi, O., Tak, P. P. & Reedquist, K. A. Histone deacetylase inhibitors suppress rheumatoid arthritis fibroblast-like synoviocyte and macrophage IL-6 production by accelerating mRNA decay. Ann. Rheum. Dis. 71, 424–431 (2012).
-
Hull, E. E., Montgomery, M. R. & Leyva, K. J. HDAC inhibitors as epigenetic regulators of the immune system: Impacts on cancer therapy and inflammatory diseases. BioMed Res. Int. 2016, 8797206 (2016).
Acknowledgements
We are grateful to our yoga therapist, Ms. Richa Mishra, for her patience and expertise in managing the patients and to all the patients who participated in this study.
Funding
We are thankful to the Department of Science and Technology (DST), India [SR/SATYAM/55/2016] for funding provided to RD. SG is supported by the Senior Research Fellowship (SRF) [45/2/2019-ANA/BMS] awarded by the Indian Council of Medical Research (ICMR), India. The funders had no role in the decision to publish or preparation of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gautam, S., Kumar, R., Kumar, U. et al. Yoga maintains Th17/Treg cell homeostasis and reduces the rate of T cell aging in rheumatoid arthritis: a randomized controlled trial.
Sci Rep 13, 14924 (2023). https://doi.org/10.1038/s41598-023-42231-w









