Dinosauria—Owen, 184225,
Ornithischia—Seeley, 188726,
Thyreophora—Nopcsa, 191527,
Jakapil kaniukura gen. et sp. nov. (Figs. 1, 2, 3, 4, Suppl. Figs. 2, 3).
Etymology
The genus, Jakapil (Ja-Kapïl: shield bearer), comes from the ‘gananah iahish’, Puelchean or northern Tehuelchean language. The specific epithet, comprising kaniu (crest) and kura (stone), refers to the diagnostic ventral crest of the mandible, and comes from the Mapudungun language. These languages, currently spoken by more than 200,000 people, have been combined as a tribute to both of the coexisting native populations of North Patagonia, South America.
Holotype
MPCA-PV-630 is a partial skeleton of a subadult individual (see Supplementary Information) that preserves fragments of some cranial bones (premaxilla, maxilla and basisphenoid), approximately 15 partial teeth and fragments, a nearly complete left lower jaw plus an isolated surangular, 12 partial vertebral elements, a complete dorsal rib and fifteen rib fragments, a partial coracoid, a nearly complete left scapula, a partial right scapula, two partial humeri, a possible partial right ulna, a complete and a partial metacarpal bone, three ischial and two femoral fragments, the distal end of a right tibia, the proximal end of a right fibula, three pedal phalanges, and more than forty osteoderms.
Referred specimens
MPCA-PV-371, two partial conical osteoderms.
Locality and horizon
Upper beds of the Candeleros Formation, early Late Cretaceous (Cenomanian, ~ 94–97 My, see16, and references therein), locality of Cerro Policía, Río Negro Province, North Patagonia, Argentina (Suppl. Fig. 1).
Diagnosis
Jakapil differs from all other thyreophorans in having: a large, ventral crest on the posterior half of the lower jaw, which is composed of the dentary, the angular and the splenial (medially hidden by the crest); a dorsomedially directed process in the short retroarticular process; leaf-shaped tooth crowns with a prominent mesial edge on their labial surface; maxillary and dentary tooth crowns differ from each other in their apical contour, the former being pointed and strongly asymmetrical, and the latter slightly curved distally with a more rounded and less asymmetrical contour; elongated (articular surface almost or completely beyond the posterior centrum face) and slender (width of less than a half postzygapophyses length) postzygapophyses in dorsal vertebrae; a strongly reduced humerus relative to the femur (proximal humeral width smaller than distal femoral width, see Supplementary Information), with a deep proximal fossa distally delimited by a curved ridge; a very large fibula relative to the femur (anteroposterior length of the proximal end almost comparable to the distal width of the femur); flattened and thin disk-like postcranial osteoderms.
Summarized description
A detailed description of the holotype is provided in the Supplementary Information. Jakapil is a small thyreophoran dinosaur (the subadult holotype is estimated to have been less than 1.5 m in body length and to have weighed 4.5–7 kg; see Supplementary Information, femoral description), with several novelties for a thyreophoran dinosaur.
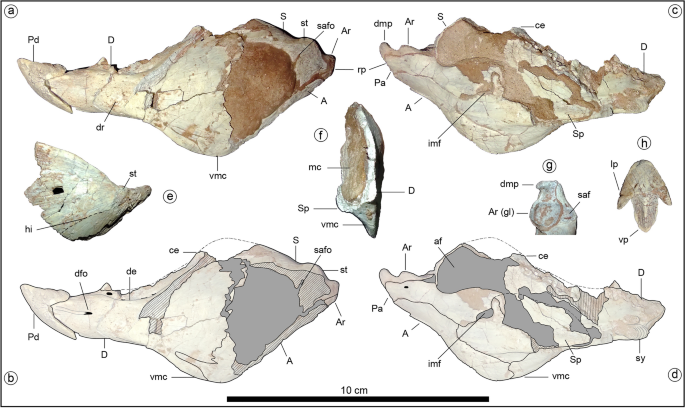
A short skull is suggested by the size of the skull and jaw bones, and the reduced number of dentary tooth positions (eleven), compared with most non-ankylosaurid thyreophorans28,29. The antorbital and mandibular fenestrae seem absent, as in ankylosaurs29 (Fig. 1a; the mandibular fenestra is also absent in Scelidosaurus10). Dentary and maxillary emarginations are present, as usual in ornithischians30 (Fig. 1a). The block-like basisphenoid is strongly similar to that of Scelidosaurus10, with Vidian canals opened posterodorsally to the basipterygoid processes, the basipterygoid processes lateroventrally projected (unlike the anteriorly directed processes of stegosaurs28 and ankylosaurs29), and a strong cultriform process (as in Lesothosaurus31, Thescelosaurus32 and probably Scelidosaurus10; Fig. 1b).
Jakapil also bears the first predentary bone (Fig. 2a–d) with a plesiomorphic shape in a thyreophoran. It is subtriangular and quite similar to that of Lesothosaurus31, and externally it is ornamented by sulci and foramina, suggesting the presence of a keratinous beak. A beak is also supported in the edentulous and subtly ornamented preserved part of the premaxilla, as in derived thyreophorans28,29. The posterior half of the short lower jaw (Fig. 2a–f) is strongly dorsoventrally expanded, resembling the general shape of the heterodontosaurid33 and basal ceratopsian jaws34. This expansion is composed of a well-developed coronoid eminence (Fig. 2a–d, ce; similar to that in the stegosaur Huayangosaurus35 and most ankylosaurs36) and a large ventral crest at the dentary-angular contact that is unique among thyreophorans (Fig. 2a–d,f, vmc; resembling that of some ceratopsians, see SI). The dentary symphysis is slightly spout-shaped, as in most ornithischians37. Anteriorly, the dentary oral margin is subhorizontal in lateral view (Fig. 2a–d, D), unlike the strongly downturned line of most thyreophorans30,37. There is no evidence of a mandibular osteoderm as occurs in Scelidosaurus and ankylosaurs10. A surangular tubercle (Fig. 2a, st) adjacent to the glenoid fossa seems anteriorly continued by a subtly developed subhorizontal inflection of the anterior lamina (Fig. 2e, hi), in the position of the surangular ridge (synapomorphy of Thyreophora37), though the first is poorly developed. The glenoid fossa is roughly aligned with the tooth row in lateral view (Fig. 2a–d). The short retroarticular process bears a dorsomedially directed process resembling that of several theropods (Fig. 2g, dmp; see Discussion). This process is absent in all other thyreophorans 9,10,35,36.
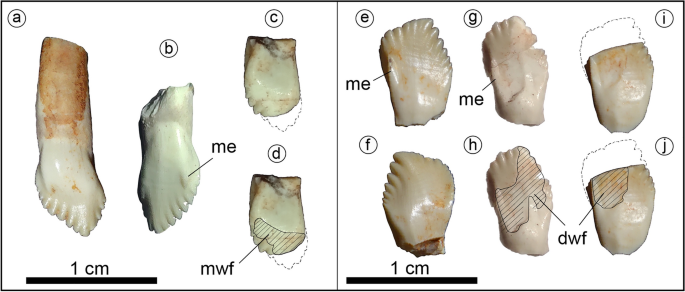
The tooth crowns are leaf-shaped as in basal ornithischian and thyreophorans10,28,29,38 (Fig. 3). The tooth crowns are swollen labially at their base and lack both cingulum and ornamentation, unlike those of derived eurypodans28,29, heterodontosaurids33 and most neornithischians30,32. The mesial edge of the labial surface in the maxillary and dentary tooth crowns is prominent as in Scelidosaurus10, and ends distally in a denticle-like structure in Jakapil (Fig. 3, me). This prominent edge delimits anteriorly the wear facets of the dentary teeth. A striking difference with respect to most thyreophorans is that the maxillary and dentary tooth crowns are quite different (see Supplementary Information). The maxillary teeth (Fig. 3a–d) show seven/eight mesial and four distal denticles, a vertical apical denticle, and a straighter mesial denticle row (resembling those of non-ankylosaurid and non-stegosaurid thyreophorans10,35,36). The dentary teeth (Fig. 3e–j) bear seven mesial and five/six distal denticles, and a distally curved apical-most denticle. Also, the mesial denticle row is lingually recurved, as in Huayangosaurus35. Large, high-angled wear facets are present (Fig. 3d,h,j; dwf and mwf).
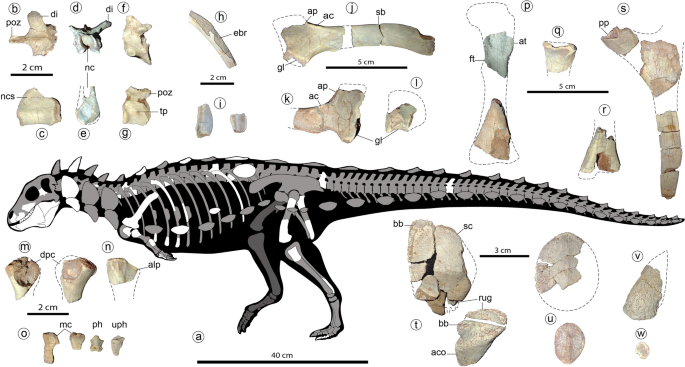
The axial elements are similar to those of Scelidosaurus39 (Fig. 4). The posterior articular surface of an isolated cervical centrum is flattened and seems almost as wide as high. A large foramen is placed just posteroventral to the parapophysis. The dorsal centra are cylindrical and elongated, with subcircular articular surfaces, and are biconcave (Fig. 4c,e). The neural arch is low but the neural canal is larger (Fig. 4d,e, nc). A dorsal neurocentral suture is visible (Fig. 4c, ncs). The diapophyses are laterodorsally directed almost 40° from the horizontal (Fig. 4d, di), at a lower angle than in stegosaurs28 and most ankylosaurs29, unlike the horizontal processes of basal ornithischians38. The postzygapophyses are medially fused in a slender (width of less than a half postzygapophyses length) and strongly elongated posteriorly structure (Fig. 4b, poz; more than in some ankylosaurs, such as Euoplocephalus and Polacanthus; see40,41). An isolated mid-caudal vertebra shows an equidimensional centrum in lateral view, with concave, oval articular surfaces (Fig. 4g). Transverse processes are very small and button-like (Fig. 4g, tp). Postzygapophyses are medially fused and do not extend beyond the centrum edge (Fig. 4g, poz). Proximally, the cross-section of the dorsal ribs is T-shaped. The low curvature of the shaft suggests a wide torso, as occurs in Emausaurus42, Scelidosaurus39, and ankylosaurs29. Some rib fragments with expanded (though broken) posterior edges suggest the presence of intercostal bones (Fig. 4h, ebr), as in Scelidosaurus39, Huayangosaurus43,44, some ankylosaurids45 (and references therein) and some basal ornithopods46. Some ribs are distally expanded (Fig. 4i) like the anterior dorsal ribs of Scelidosaurus39 and Huayangosaurus43.
Girdle and limb bones (see also Suppl. Figs. 2, 3) are mostly broken and with boreholes (probably due to bioerosion) at their ends. The scapular blade (Fig. 4j, sb) is elongated and parallel-sided, without distal expansion, an overall shape that resembles that of several theropods47, contrasting the distally expanded condition in most ornithischians30. A straight and parallel sided scapular blade is common in ankylosaurids29,40. The proximal scapular plate with a high acromial process (Fig. 4j,k, ap) is stegosaurian-like, and the lateral acromial crest (Fig. 4j,k, ac) is developed as in Huayangosaurus43. A low distinct ridge rises posterior to the glenoid fossa and represents the insertion site for the muscle triceps longus caudalis, as occur in ankylosaurids 40. The incomplete coracoid (Fig. 4l) is much shorter than the scapula, unlike that of ankylosaurs29,40, which bear a large coracoid. The coracoid and the scapula are not fused. The partial humeri (Fig. 3m) are strongly reduced in size, with overall limb proportions resembling those of basal ornithischians3,38 and several theropods47. A possible proximal end of the ulna (Fig. 4n) resembles that of other basal ornithischians, though more strongly laterally compressed. The anterolateral process is present (Fig. 4n, alp), and the olecranon process seems absent or poorly developed, as in Scutellosaurus9 and Scelidosaurus39. The ischia are poorly preserved (Fig. 4s). The pubic peduncle is separated from the iliac articulation, unlike the continuous cup-shaped structure of most ankylosaurs29. The shaft of the ischium is straight and parallel-edged, as in Scutellosaurus9 and Scelidosaurus39, and distally tapers as in stegosaurs28. The preserved femoral pieces (Fig. 4p) resemble those of basal ornithischians38,39. The bases of both the broken anterior and fourth trochanters (Fig. 4p, at, ft) are large, suggesting large elements; the fourth trochanter is proximally placed on the femoral shaft (near the height of the base of the anterior trochanter); and the distal end of the femur is slightly curved posteriorly. The proximal end of the right fibula (Fig. 4q) is much larger than that of all other thyreophorans (compared with both the femoral and tibial distal ends) and bears a large anterior curved crest. The block-like non-ungual phalanges and a bluntly pointed hoof-like ungual (Fig. 4o, ph, uph) are similar to those of Scelidosaurus39.
At least five osteoderm types are preserved in the holotype of Jakapil. The cervical elements are composed of an external, low-crested scute (Fig. 4t, sc) over a fused, smooth bone base (Fig. 4t, bb), as in Scelidosaurus48 and several ankylosaurs2,49. A probable cervical element is also composed of a concave base of smooth bone fused to a high, asymmetrical osteoderm (Fig. 4t, aco). The bases of these dermal elements present strong rugosities at one edge, suggesting a sutural contact between (Fig. 4t, rug), as in Scelidosaurus48 and some ankylosaurs (such as Pinacosaurus and Scolosaurus40,49,50). Scute-like post-cervical osteoderms (Fig. 4u) are strongly flattened, disk-shaped, and suboval with a very low crest, resembling those of few ankylosaurs such as Gastonia and Gargoyleosaurus51 (‘body osteoderms’ sensu Kinneer et al.52; see also49). Only one scute shows a high triangular cross-section like those of Scelidosaurus48. Also present are a few conical, spike-like osteoderms with deep concave bases (Fig. 4v), and many flat, disk-shaped, minute (7–10 mm) ossicles without crests (Fig. 4w).
Phylogeny
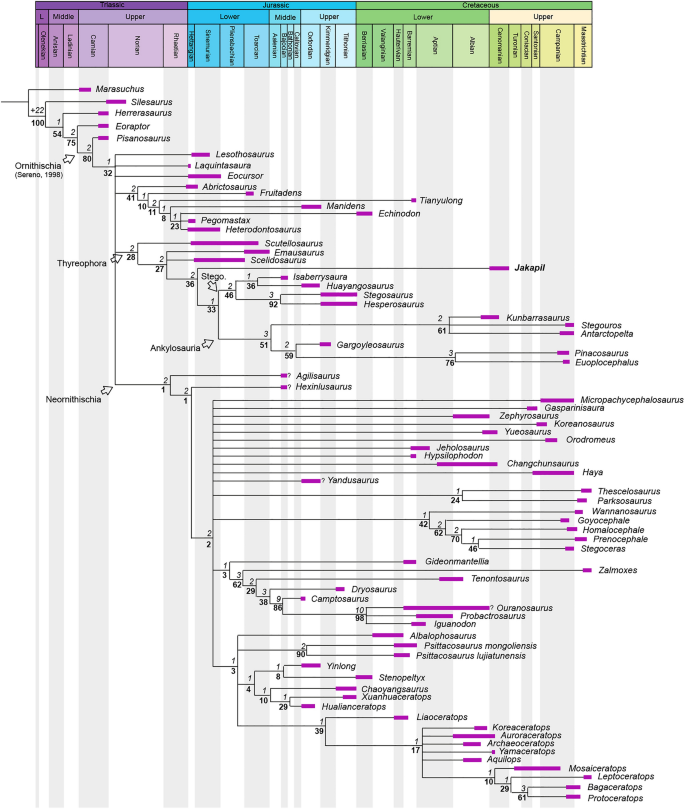
The phylogenetic analysis using the matrix of Soto-Acuña et al.5 recovers Jakapil within Thyreophora, as the sister taxon of Ankylosauria (Fig. 5). The branch support for the basal thyreophorans is considerably lower than that obtained by Soto-Acuña et al.5, although the support of Stegosauria and some less inclusive eurypodan clades is slightly better (ceratopsians and pachycephalosaurs also show a lower support). The Jakapil autapomorphies in this analysis are: ventrally orientated basipterygoid processes (char. 134; shared with Agilisaurus, Hypsilophodon, Zalmoxes, Tenontosaurus, Dryosaurus, Liaoceratops, Yamaceratops, Leptoceratops, Bagaceratops and Protoceratops); lateral orientation of the basipterygoid process articular facet (char. 136; shared with Homalocephale, Prenocephale, Stegoceras and Yinlong); a straight dentary tooth row in lateral view (char. 166; shared with the ornithischians Lesothosaurus, Eocursor, Scutellosaurus, Pinacosaurus, Euoplocephalus, heterodontosaurids and neornithischians); the presence of a ventral flange on the dentary (char. 170; shared with Psittacosaurus, Yamaceratops and Protoceratops); a well-developed coronoid process (char. 174; shared with heterodontosaurids and neornithischians); a surangular length of more than 50% the mandibular length (char. 183; shared with Stegoceras, Psittacosaurus, Yinlong, Chaoyangsaurus and Hualianceratops); less than 15 dentary teeth (char. 204; shared with heterodontosaurids, Gasparinisaura, Hypsilophodon, Wannanosaurus, Tenontosaurus, Dryosaurus and ceratopsians); apicobasally tall and blade-like cheek teeth crowns (char. 205; shared with Laquintasaura, Psittacosaurus, Yinlong, Chaoyangsaurus and Hualianceratops). Alternative phylogenetic analyses using the data matrices of Maidment et al.4, Norman6 and Wiersma and Irmis8 recover Jakapil as the sister taxon of Eurypoda (Stegosauria + Ankylosauria) and as a basal ankylosaur, respectively (see Supplementary Information). Being recovered either as an ankylosauromorph or a stem-eurypodan, Jakapil is closely related to Scelidosaurus in all analyses. Detailed phylogenetic results and discussion are provided in the Supplementary Information.