









-
NASA’s Webb Telescope revealed just how giant the water plumes shooting out of a Saturn moon are.
-
The water gushes 6,000 miles, or about twice the length of the US, from the moon called Enceladus.
-
The plume comes from an underground ocean, meaning the moon could host alien life.
NASA’s James Webb Space Telescope spotted a gigantic water plume spewing from a Saturn moon that could host alien life. It’s creating a donut of water around the ringed planet.
Astronomers have previously seen water shooting from the moon, called Enceladus, but never like this.
NASA’s Cassini spacecraft first spotted the mysterious spray on the moon’s south pole as it flew past in 2005. Before that, scientists thought Enceladus was an inert ball of ice. The plumes revealed something far more exciting: a global ocean deep below its frozen crust. That makes Enceladus one of our solar system’s top candidates for alien life.
The close-up Cassini images gave astronomers this vision of Enceladus:

But the James Webb Space Telescope is the most powerful observatory ever launched into space. So when scientists turned its sites to Enceladus, they saw its salty water plume at an unprecedented scale that indicates these plumes are much larger than previously thought.
Webb revealed an enormous plume on Enceladus
Webb discovered a 6,000-mile-long burst of water — twice the length of the continental US — NASA announced on Tuesday.

“When I was looking at the data, at first, I was thinking I had to be wrong,” Geronimo Villanueva, a scientist at NASA’s Goddard Space Flight Center and a lead author on the new Webb findings, said in a press release. “It was just so shocking to detect a water plume more than 20 times the size of the moon.”
The plume is long enough to stretch from Los Angeles, California, to Buenos Aires, Argentina. For comparison, Enceladus itself could fit comfortably between Los Angeles and San Francisco.
The plumes are exploding off the moon’s surface into space at a rate of 79 gallons per second, which could fill an Olympic-sized swimming pool in just two hours, according to NASA.
Enceladus formed a water donut around Saturn
About 30% of the water from the plumes then feeds into a giant ring of water around Saturn, called a torus, and the rest escapes into the planet’s water system.
“The orbit of Enceladus around Saturn is relatively quick, just 33 hours. As it whips around Saturn, the moon and its jets are basically spitting off water, leaving a halo, almost like a donut, in its wake,” Villanueva said.
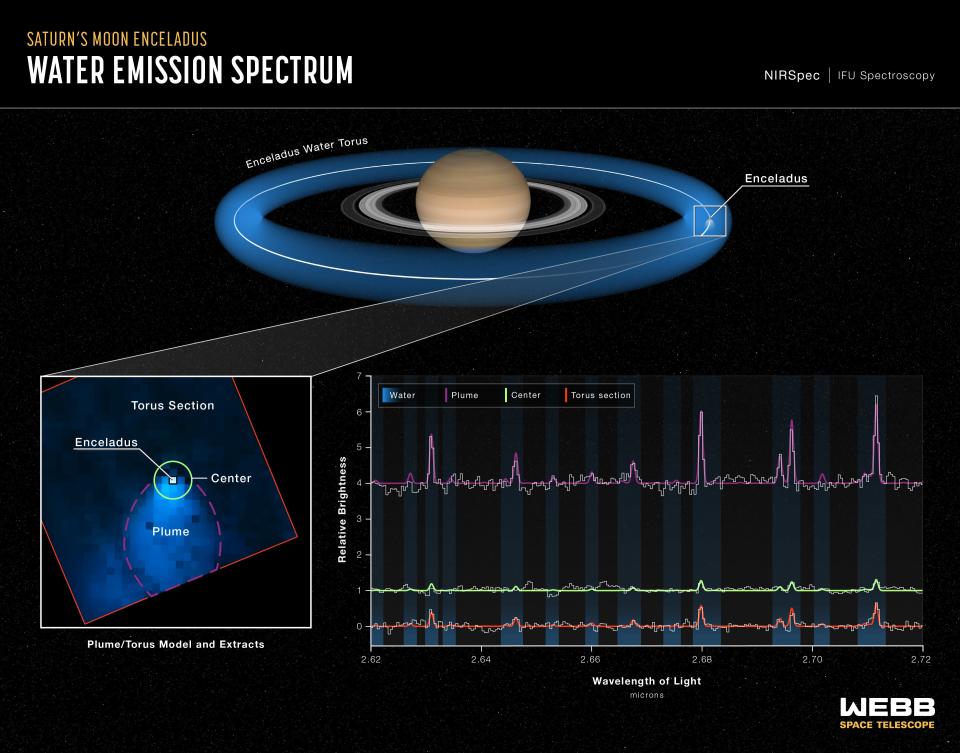
“In the Webb observations, not only was the plume huge, but there was just water absolutely everywhere,” Villanueva said.
Saturn has at least 124 moons, and Enceladus is among its most interesting.
It’s about 4% the size of Earth. Any alien life in its subsurface ocean could be getting energy from deep-sea vents, using chemosynthesis rather than photosynthesis, as some deep-sea organisms do on Earth.
Read the original article on Business Insider









