








An important system of ocean currents that circulates water around the planet could significantly slow down or even stop completely in just a few decades, according to a shocking new study released Tuesday.
The network is called the Atlantic Meridional Overturning Circulation, or AMOC, which includes the Gulf Stream. It’s a series of ocean currents that brings warm water north, and cold water south across the Atlantic Ocean, part of a “global conveyor belt” that impacts weather patterns across North America, Europe, Asia and Africa, according to NOAA.
Researchers in Denmark analyzed sea surface temperatures to determine the strength of the AMOC, using data from 1870 to 2020. The pair, Susanne Ditlevsen of the University of Copenhagen, and her brother, Peter Ditlevsen of the university’s Niels Bohr Institute, then created a statistical model to analyze early-warning signals that there are problems with the current network.
The authors concluded the AMOC could collapse at any point between now and 2095, even as early as 2025.
Their models rely on “the current scenario of future emissions,” assuming that greenhouse gases would continue to be released into the atmosphere without dramatic steps to reduce them. The new research was published Tuesday in the journal Nature Communications.
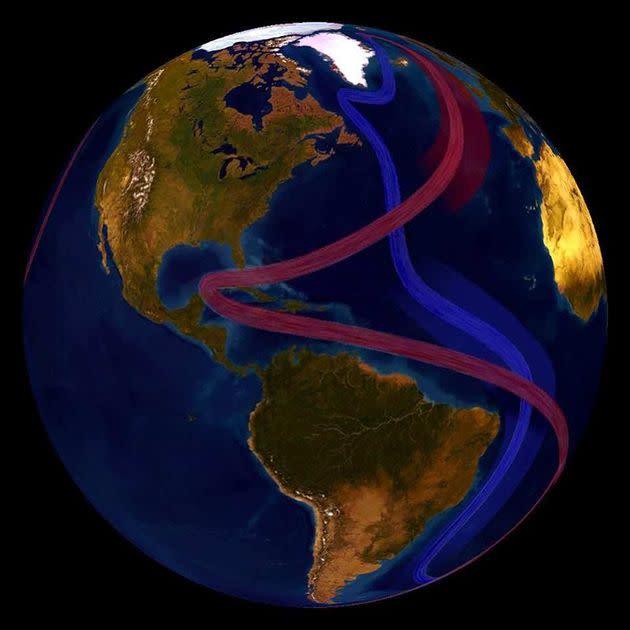
The global conveyor belt, shown in part here, circulates cool subsurface water and warm surface water throughout the world. The Atlantic Meridional Overturning Circulation, or AMOC, is part of this complex system of global ocean currents.
A collapse “would have severe impacts on the climate in the North Atlantic region,” the authors wrote, and represent one of the most important “tipping points” as the planet’s climate changes. Other tipping points — which represent irreversible shifts to the planet — include the melting of the Greenland ice sheet, the destruction of the Amazon rainforest and the thawing of the permafrost.
The Washington Post notes the analysis is different from that in the most recent climate report by the United Nations Intergovernmental Panel on Climate Change. The ICCC concluded at the time it had “medium confidence” the AMOC would not fully collapse this century.
Projects to monitor the health of the AMOC have been gathering data since 2004, but some scientists also say that the short time frame isn’t long enough to extrapolate predictions about how the ocean could change over the coming decades.
The authors of the paper also acknowledged that they could not rule out “other mechanisms are at play” in the changes to the AMOC.
Still, Michael Mann, a climate scientist at the University of Pennsylvania, told Axios that while there were some questions about the study’s results, they only added to growing concern about the state of the planet amid uncontrolled climate change.
“I think the authors in this case are on to something real,” Mann told the outlet.
The sudden shutdown of the AMOC was the key element in the 2004 disaster movie, “The Day After Tomorrow.” While the actual collapse of the current system is unlikely to produce immediate catastrophic weather changes, it could cause colder temperatures in northern Europe and warming in tropical zones, Peter Ditlevsen told the Post.
“This is a really worrying result,” he said, adding to the publication that the evidence demonstrated further need for a “hard foot on the break” of carbon emissions.
The authors said the results should call for “fast and effective measures to reduce global greenhouse gas emissions.”









