








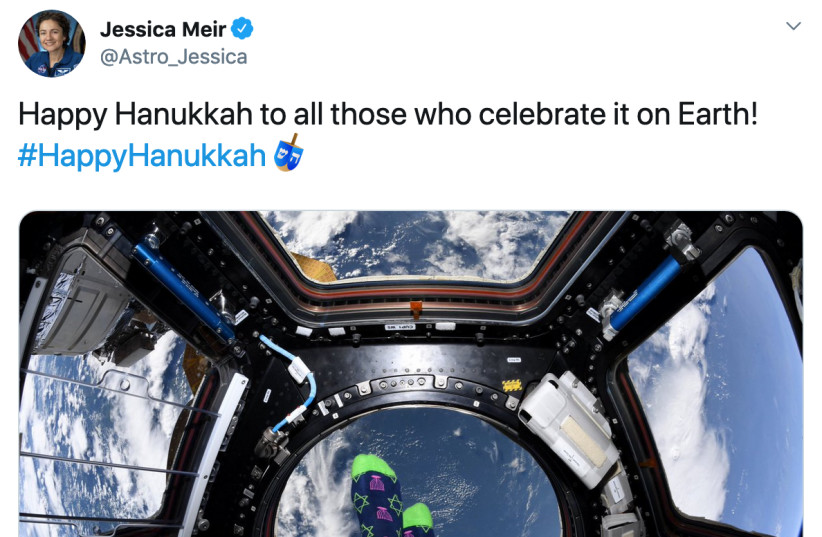
Since the holiday kicked off on Sunday, Hanukkah wishes and pictures are flooding social media, including many messages from international leaders and celebrities from all over the world. However, an exceptional Hanukkah greeting was even able to transcend the Earth’s borders.“Happy Hanukkah to all those who celebrate it on Earth!” astronaut Jessica Meir tweeted straight from the International Space Station.
— Jessica Meir (@Astro_Jessica) December 23, 2019
Meir, the daughter of an Israeli doctor who moved to Sweden and later to the US, joined the crew of the ISS in September. Two weeks later she participated in the very first all-female spacewalk.As lighting candles every night is the core tradition of the festival, Hanukkah offers plenty of opportunities for evocative pictures and compelling posts on social media (every possible variation of the theme “a light in the dark” gets explored). But lighting fires in outer space can be especially challenging, considering that flames can react to the absence of gravity in an uncontrolled manner. Perhaps for this reason, for her messageMeir opted for a picture of her feet wrapped in a pair of entertaining socks featuring pink menorahs and green stars of David on a blue background. And she did follow the tradition of placing the menorah close to the window, since behind her feet, the earth is visible thousands of kilometers away. One question stands: did Meir bring the designed socks from the earth when she left several months ago or did she get them delivered to space with one of the regular shipments to the ISS? The bottom line is: it’s never too early to get ready for the holidays.









